Embriologia del pollo
Anatomia
funzionale degli uccelli domestici
Virgilio BOTTE - Gaetano PELAGALLI
Edi-Ermes, Milano, 1982

Il frequente uso delle uova embrionate di uccelli per indagini microbiologiche rende utile la descrizione degli annessi embrionali, uno dei quali, la membrana corioallantoidea, costituisce il tessuto di elezione per colture in vivo di un'ampia varietŕ di agenti patogeni.
Nei vertebrati ovipari inferiori, come gran parte dei selaci (i pesci cartilaginei), dei pesci ossei e degli anfibi, lo sviluppo embrionale deve avvenire necessariamente in acqua (dolce o di mare) in quanto le loro uova, se deposte in ambiente asciutto, non riescono a evitare un rapido essiccamento. Tale limite č stato superato con la comparsa dei rettili, precursori degli uccelli, nel cui uovo fecondato sin dai primi stadi di accrescimento dell'embrione si forma una membrana, l'amnios[1], che crea intorno al germe una cameretta umida, quasi un lago in miniatura. Questa importante acquisizione, associata ad altre giŕ presenti nei vertebrati meno evoluti, come il sacco del tuorlo, e la messa a punto di una terza struttura capace di operare gli scambi gassosi con l'ambiente, l'allantoide[2], hanno rappresentato il presupposto per lo sviluppo lontano dall'acqua e quindi hanno permesso ai rettili di invadere la terraferma.
Per comprendere l'organizzazione e le tappe della formazione delle membrane prima ricordate, spesso indicate nel loro complesso come annessi embrionali, sarŕ utile seguire nelle linee generali gli stadi precoci dello sviluppo embrionale, rinviando a testi specializzati per maggiori dettagli.
Formazione della discoblastula
Nelle uova di uccelli la ricchezza di vitello condiziona la segmentazione dell'uovo fecondato, che viene a essere praticamente confinata al polo animale ove viene a trovarsi il nucleo con una scarsa quantitŕ di citoplasma. La meiosi nell'ovocita si compie in parte durante l'ultima fase dell'accrescimento nell'ovario e in parte nella regione piů alta dell'ovidutto dopo la penetrazione di piů spermatozoi (polispermia). Uno solo di questi, comunque, si fonde con il nucleo femminile a costituire lo zigote, dalla cui divisione inizia lo sviluppo embrionale.
In caso di fecondazione, le divisioni dello zigote durante il tragitto nell'ovidutto si susseguono, ma non interessano tutto l'uovo, perché impedite dall'enorme massa di vitello. Alla deposizione dell'uovo, sul polo animale «galleggia» un piccolo gruppo di cellule, appena visibile a occhio nudo, che rappresenta la discoblastula. L'esame istologico permette di stabilire che in essa sono presenti due componenti: uno superficiale, l'ectoblasto, e l'altro profondo, l'endoblasto.
Il brusco abbassamento della temperatura dopo la deposizione blocca ogni ulteriore sviluppo del germe, che potrŕ riprendere quando l'uovo verrŕ messo in un ambiente a temperatura adeguata e caratteristica a seconda della specie.

Fig.
1
1
- Aree organoformative alla superficie del disco germinativo di un uccello.
2 a - Disco germinativo di uccello visto in superficie.
2 b - Disco germinativo di uccello visto in sezione, secondo il piano passante
per la linea S.
(da Williams & Wendel-Smith, Istituzioni di Embriologia, Edi. Ermes)
Gastrulazione
Alla deposizione dell'uovo, nel pollo la discoblastula ha un diametro medio di circa 4 mm e appare distinta in due porzioni: centrale piů trasparente, l'area pellucida, e periferica piů estesa, l'area opaca. La prima darŕ origine all'embrione, mentre la seconda rappresenterŕ il materiale da cui si svilupperanno gli annessi embrionali (Fig.1).
Agli inizi dell'incubazione i margini periferici della discoblastula si espandono per rivestire il tuorlo, mentre nell'area pellucida avviene la gastrulazione. Questo processo ha lo scopo di portare nella sua giusta posizione il materiale destinato a dare il mesoderma e la corda dorsale, che si trova compreso nell'ectoblasto.
Come č noto, il mesoderma deve venire a trovarsi tra ectoderma ed endoderma, e la corda dorsale deve occupare una posizione mediana sagittale, secondo l'asse lungo del futuro embrione, inferiormente al tubo neurale e superiormente al canale alimentare. Gli opportuni spostamenti dei tessuti embrionali si realizzano mediante scivolamento lungo un'area di invaginazione che corre secondo l'asse lungo del futuro embrione, la linea primitiva (Fig. 2).
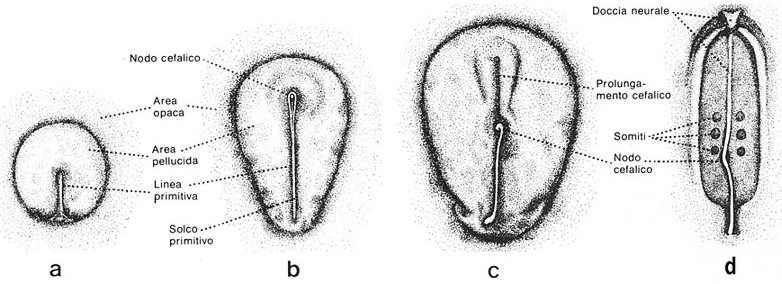
Fig.
2
Varie
fasi di sviluppo di un embrione di uccello, visione in superficie.
(da Williams & Wendel-Smith, Istituzioni di Embriologia, Edi. Ermes)
Al termine della gastrulazione, l'embrione appare formato da tre strati di tessuto: il superficiale porta su una fascia longitudinale mediana il neuroectoderma, futuro tubo neurale, e, intorno a questo, l'ectoderma; il medio č costituito dalla corda dorsale, mediana, fiancheggiata dal mesoderma; l'interno č fatto dall'endoderma, che fornirŕ gran parte del canale alimentare. In questo stadio i tre strati sono aperti ventralmente e galleggiano sul sottostante vitello.
Il mesoderma č suddivisibile in varie regioni che, da ciascun lato, procedendo dalla corda dorsale alla periferia, sono: i somiti o mesomeri[3], metamerici (metameri = serie di concamerazioni appaiate lungo l'asse longitudinale), e la piastra laterale. Quest'ultima appare presto distinta in una lamina superficiale, la somatopleura, accollata all'ectoderma, e in una profonda, la splancnopleura, aderente all'endoderma. Tra le due lamine viene a definirsi una cavitŕ: il celoma.
Le lamine mesodermiche invadono anche l'area opaca, a sua volta distinta in una parte piů prossima all'area pellucida, l'area vascolosa, e una piů periferica, l'area vitellina. La prima zona č particolarmente importante perché dal suo mesoderma si sviluppano i vasi sanguigni e il sangue, che diffonderanno negli annessi embrionali e si congiungeranno ai vasi frattanto formatisi nell'embrione.
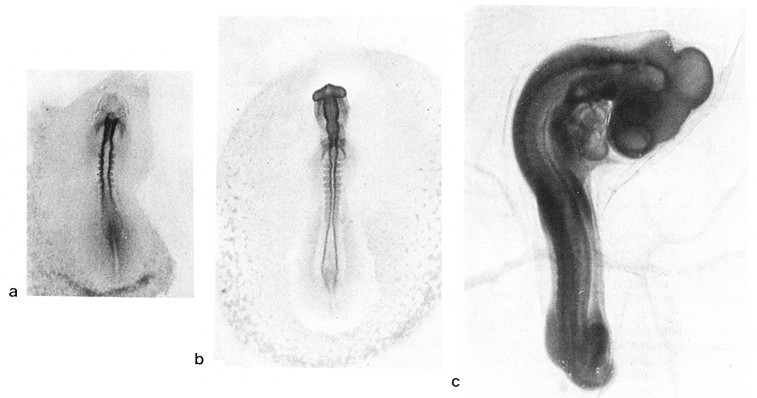
Fig.
3
Immagini
di embrioni di pollo, ai primi stadi di sviluppo.
a - 24 ore di sviluppo; b - 33 ore; c - 72 ore.
Sollevamento dell'embrione
Quando la gastrulazione č compiuta, le aree che daranno l'embrione si definiscono dalle circostanti a opera di pliche, delle quali la piů precoce č la plica cefalica, posta appena innanzi alla regione che svilupperŕ il capo. L'embrione progressivamente si solleva e la sua continuitŕ con le regioni extraembrionali si riduce a una ristretta area ventrale, l'ombelico, da cui parte il cordone ombelicale che continua negli annessi embrionali. In questo stesso periodo, lungo la linea sagittale dorsale dell'embrione, il neuroectoderma si chiude nel tubo neurale e l'ectoderma passa a rivestirlo esternamente. La tipica organizzazione del vertebrato č ormai acquisita (Fig. 3).
Sviluppo degli annessi embrionali
L'area vitellina appare distinta in una zona superficiale o esterna e una profonda o interna. La prima č costituita da ectoderma e somatopleura, la seconda da endoderma e splancnopleura. Tra i due foglietti viene a trovarsi, ovviamente, la cavitŕ celomatica che, dopo il sollevamento dell'embrione, si segrega dal celoma embrionale e perciň viene indicata come celoma extraembrionale.
Sacco del tuorlo — Mentre nell'area pellucida avvengono i fenomeni prima descritti, la discoblastula dell'area opaca non ancora raggiunta dal mesoderma procede nel rivestire il tuorlo. Nel pollo, alla fine del secondo giorno di incubazione, ha raggiunto l'equatore e il processo č pressoché completo al 6° giorno (resta una piccola area scoperta, l'ombelico ombelicale, che scomparirŕ al 16° giorno). Il vitello viene avvolto, cosě, dalla lamina endodermica, in continuitŕ con quella intestinale embrionale attraverso un breve canale vitellino; al suo esterno si appone la splancnopleura, la quale porta i vasi sanguigni, sacco del tuorlo. Il tuorlo puň cosi essere riassorbito e trasferito per via sanguigna all'embrione. Con la diminuzione del vitello, il sacco del tuorlo si riduce progressivamente; alla nascita sono presenti solo suoi residui che saranno presto riassorbiti.
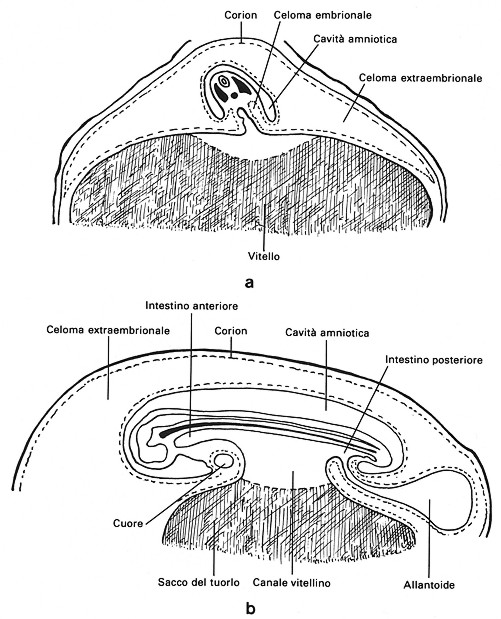
Fig.
4
Cavitŕ
amniotica e allantoide dell'embrione di pollo.
a - Sezione trasversale di un embrione al termine del secondo giorno
d'incubazione.
b - Sezione sagittale di un embrione in cui le pieghe amniotiche si sono
congiunte.
Amnios e corion — Nel pollo, verso la 30a ora d'incubazione, dalla membrana esterna dell'area vitellina si solleva una piega amniotica anteriore che, portandosi posteriormente, include la testa come in un cappuccio. All'inizio del 3° giorno una formazione analoga, piega amniotica posteriore, si accresce posteriormente raggiungendo, verso la 62a ora, l'anteriore. Resta una piccola zona non ricoperta, il poro corioamniotico, che si oblitera all'inizio del 4° giorno (Fig. 4). Al termine di questo processo l'embrione viene a essere compreso nella cavitŕ amniotica piena di fluido salino derivato dall'albume compreso tra le pliche. La parete della cavitŕ o amnios sarŕ costituita internamente da ectoderma e all'esterno da somatopleura e non č mai vascolarizzata. La parte esterna delle pieghe amniotiche formerŕ il corion o sierosa che si spinge alla periferia tendendo all'accollamento alle membrane testacee. Quest'ultimo fenomeno si realizza progressivamente con la diminuzione dell'albume. La principale funzione della cavitŕ amniotica č quella di consentire lo sviluppo dell'embrione in ambiente umido. Il liquido amniotico č sottoposto a continui movimenti dalle fibrocellule muscolari differenziate nella somatopleura; come nei mammiferi, viene anche rinnovato. L'amnios, indirettamente, provvede alla digestione dell'albume che tra l'11° e il 16° giorno d'incubazione puň penetrare nella sua cavitŕ attraverso una piccola discontinuitŕ ed essere inghiottito dall'embrione. Al 16° giorno la comunicazione si oblitera. Alla schiusa l'embrione inghiotte tutto il liquido amniotico; l'amnios resta legato al guscio (Fig. 5).
Allantoide — Č un diverticolo ventrale della porzione posteriore dell'intestino costituito da endoderma rivestito della splancnopleura. Inizia il suo sviluppo alla 60a ora d'incubazione e si proietta in avanti portandosi entro il celoma extraembrionale. L'embrione č inclinato sul lato sinistro, per cui l'allantoide si espande nella parte destra del celoma, viene in contatto con il corion e poi si insinua tra amnios e corion portandosi anche a sinistra. Il contatto con l'allantoide costringe il corion contro le membrane testacee e l'albume viene a essere concentrato in un sacco dell'albume posto all'apice acuto dell'uovo. A partire dal 4° giorno nella splancnopleura dell'allantoide si formano i vasi sanguigni, vasi allantoidei. La componente mesodermica si fonde presto con quella del corion che ne viene rapidamente vascolarizzato. Si costituisce cosi l'allantocorion o membrana corioallantoidea. L'allantoide esplica funzioni respiratorie di primaria importanza sino al 19° giorno, quando inizia la circolazione polmonare. Partecipa, altresě, all'assorbimento dell'acqua e di parte delle proteine dell'albume. Verso il termine del periodo d'incubazione procede anche a riassorbire parte dei sali minerali del guscio che l'embrione utilizza per il suo scheletro. Nella cavitŕ allantoidea si accumulano i prodotti di rifiuto azotato in forma di acido urico. L'acqua necessaria a disciogliere questo catabolita viene in gran parte riassorbita, per cui alla schiusa si trovano nella cavitŕ solo cristalli di urati.
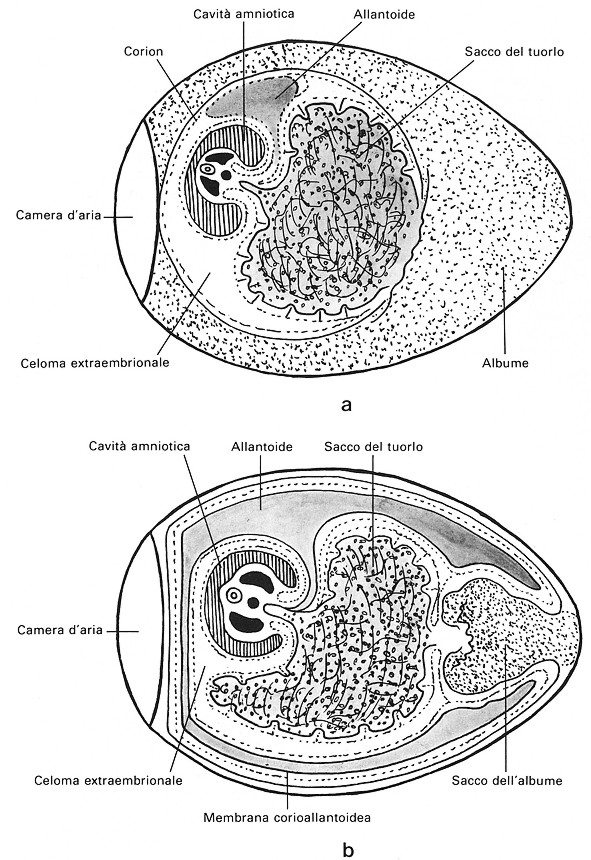
Fig.
5
Sezioni
sagittali di uova di polio al 6° (a) e al 14° giorno (b) d'incubazione.
![]()
[1] Amnio, amnion o amnios: dal greco amníon, vaso in cui si raccoglieva il sangue delle vittime. Annesso embrionale costituito da un sacco che si sviluppa a spese di una parte dei tessuti formati dall'uovo fecondato (sacco amniotico), contenente – in quantitŕ variabile nei diversi animali e nei vari stadi di sviluppo – un liquido sieroso (liquido amniotico) nel quale č immerso l'embrione ancorato al cordone ombelicale.
[2] Allantoide: dal greco allantoeidës, che ha la forma di salsiccia, essendo allâs la salsiccia, il sanguinaccio.
[3] In embriologia i mesomeri, o somiti, sono masse simmetriche del mesoderma assiale che si formano nell'uovo dopo la prima fase di formazione dei tre foglietti embrionali (endoderma, mesoderma, ectoderma). I mesomeri sono suddivisibili in tre regioni: dermatomo, da cui si origina il derma, il miotomo, da cui si origina la muscolatura, e infine lo sclerotomo, da cui si origina la colonna vertebrale e la regione occipitale del cranio.