Vol. 2░ - III.1.
ANATOMIA DELLA CELLULA EUCARIOTICA
La cellula eucariotica si compone di un protoplasma, formato da nucleo e citoplasma, e di una membrana plasmatica, che costituisce una struttura delimitante dello spessore medio di 8,5 nm e che varia a seconda del tipo di cellula.
La membrana Ŕ ricoperta dal cell
coat, il mantello cellulare, detto anche glicocalice in quanto costituito
da glicoproteine, coltre esterna morbida e flessibile dotata di spiccate
proprietÓ adesive e che al microscopio elettronico ha lĺaspetto di lanugine
sfilacciata.
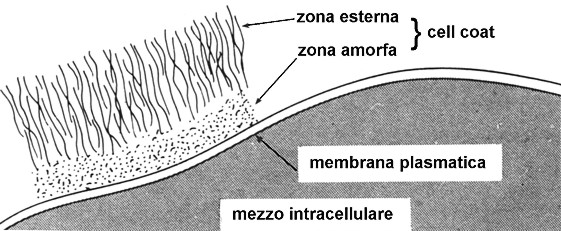
Fig. III. 2 - Membrana plasmatica e cell coat come si presentano nellĺameba
1.1. Membrana plasmatica
La membrana plasmatica Ŕ sede di numerose attivitÓ della cellula: risposta agli ormoni, interazioni cellulari, proprietÓ antigeniche, eccitabilitÓ e, forse, anche espressione dei geni e della divisione cellulare.
Allĺinizio degli anni ĺ70 Ŕ stato proposto un modello della struttura della membrana plasmatica detto modello trilaminare a mosaico, valido anche per le membrane interne alla cellula, secondo il quale in un bilayer fosfolipidico sono affondate proteine che attraversano tutta la membrana da parte a parte. I principali fosfolipidi di membrana sono sostanzialmente 4: fosfatidilserina, fosfatidiletanolamina, fosfatidilcolina e sfingomielina.
Sono molecole con unĺestremitÓ idrofoba e una idrofila, quindi anfipatiche,
che quando si trovano in mezzo a due soluzioni acquose, come lĺinterno e lĺesterno
della cellula, interagiscono tra loro disponendosi in doppio strato, strato
lipidico bimolecolare o bilayer dello spessore di circa 7-8 nm.
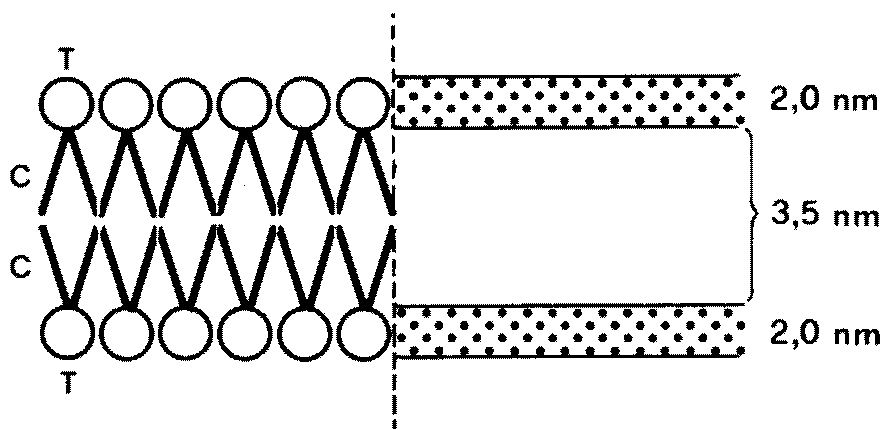
Fig. III. 3 - Membrana plasmatica in microscopia elettronica a trasmissione.
Le aree punteggiate dello spessore di circa 2 nm sono dovute alla disposizione
allineata delle teste dei fosfolipidi (T). Nello spazio chiaro interposto tra le due superfici
si giustappongono le code dei fosfolipidi (C).
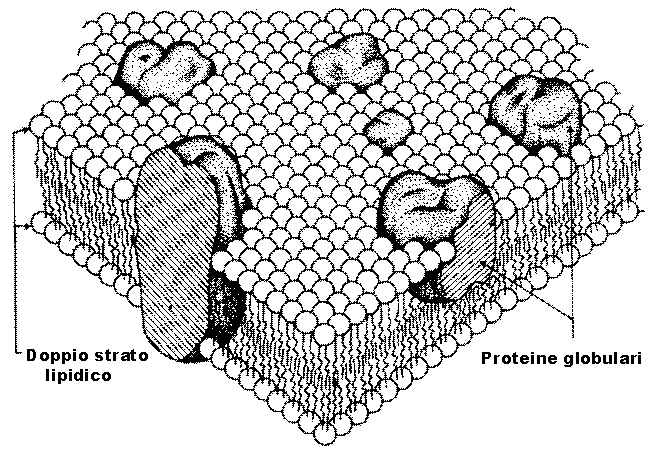
Fig.
III. 4 - Membrana plasmatica senza cell coat.
Si tratta del modello a mosaico proposto da Singer
e Nicolson.
1.2. Nucleo
Il nucleo, rotondeggiante oppure ovoidale, in seno al
nucleoplasma contiene uno o due nucleoli e zolle di cromatina.
Lĺinvolucro nucleare
Ŕ costituito da due membrane: una esterna in contatto col citoplasma e che
rappresenta la faccia citoplasmatica, una interna in rapporto con la cromatina
che vi aderisce. Uno spazio perinucleare separa le due membrane.
I pori nucleari, che si aprono attraverso lĺinvolucro nucleare,
offrono alle sostanze esogene o endogene la possibilitÓ di transitare sia in
direzione nucleocitoplasmatica che in direzione inversa.
La cromatina, materiale intensamente colorabile con coloranti
basici, Ŕ composta principalmente da nucleoproteine; essa si organizza in
filamenti pi¨ o meno spiralizzati, con diametro pari o inferiore a 25 nm.
Il nucleolo, con la sua matrice densa, appare come una rete di
maglie anastomizzate sulle quali spiccano componenti ribonucleoproteici
granulari e fibrillari disposti senza regola; occupa una zona variabile del
nucleo.
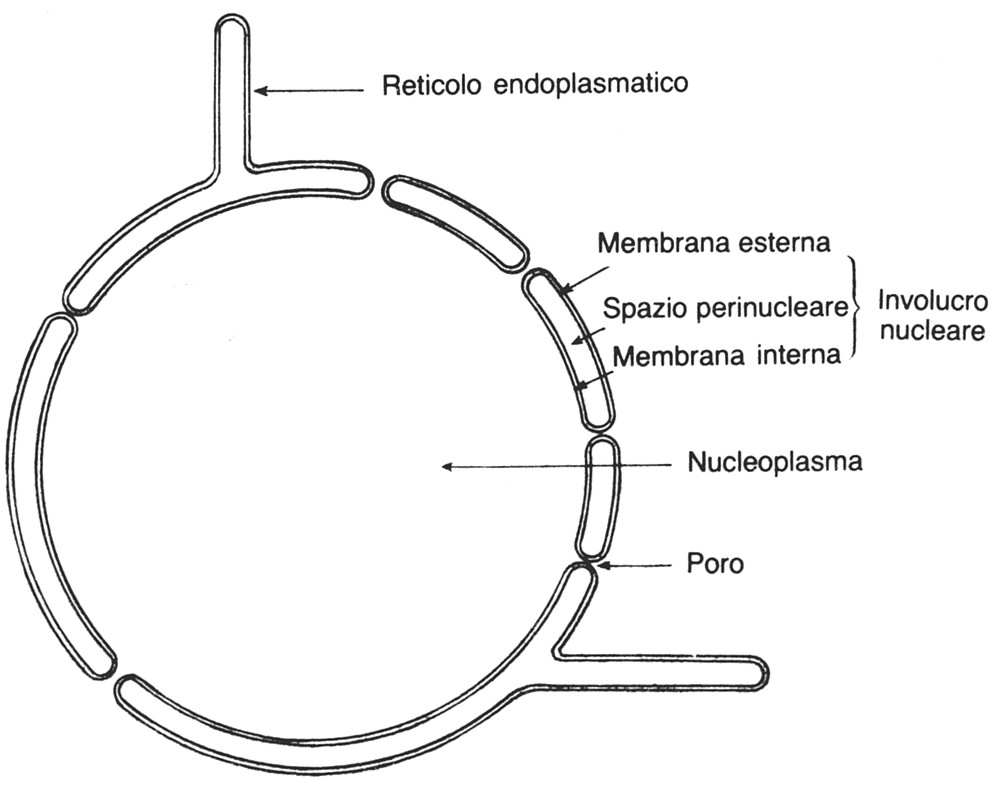
Fig. III. 5 - Lĺinvolucro nucleare
1.3. Citoplasma
Il citoplasma Ŕ costituito due
entitÓ diverse: morfoplasma e ialoplasma.
1.3.a. Morfoplasma
Il morfoplasma comprende tutti
gli elementi figurati del citoplasma, cioŔ tutti i seguenti organuli
cellulari:
Ě
reticolo endoplasmatico granulare
- REG: Ŕ lĺinsieme delle cisterne appiattite le cui
membrane, sulla faccia esterna, sono dotate di granuli detti ribosomi.
Ě
reticolo endoplasmatico liscio
- REL: Ŕ un insieme di tubuli le cui membrane sono prive di
ribosomi.
Ě
mitocondri: elementi di piccole dimensioni
con diametro medio di 1mm.
Sono limitati da due membrane: una membrana esterna liscia; una membrana
interna provvista di creste che ne aumentano la superficie.
Ě
apparato di Golgi:
rappresenta lĺinsieme dei dittiosomi
della cellula. Ciascun dittiosoma comprende cisterne impilate, vescicole di
piccole dimensioni e vacuoli disposti su una delle facce del dittiosoma quando
esso Ŕ attivo.
Ě
lisosomi: organuli delimitati da una
membrana semplice, contenenti una sostanza omogenea, moderatamente densa, le
cui molteplici attivitÓ enzimatiche possono distruggere elementi estranei
alla cellula.
Ě
perossisomi: hanno diametro simile a quello
dei lisosomi e racchiudono nella loro membrana un nucleoide (formazione
paracristallina) immerso in una sostanza omogenea.
Ě
centrosoma: si trova in genere vicino al
nucleo. ╚ formato da uno o due centrioli. Ciascun centriolo ha lĺaspetto di un cilindro, la cui parete
leggermente opaca contiene 9 triplette di tubuli, disposte parallelamente.
Ě
microtubuli e microfibrille,
formanti il cosiddetto citoscheletro:
isolati o raggruppati in fasci, dispersi o localizzati, percorrono il
citoplasma. Fanno parte del citoscheletro: i microfilamenti di actina e di
miosina, i filamenti intermedi e la rete microtrabecolare.
1.3.b. Ialoplasma o Citosol
Detto anche plasma amorfo, Ŕ
una soluzione dotata di pH 7, omogenea, senza strutture, che costituisce
generalmente il 55% dellĺintero volume cellulare. ╚ quella parte del
citoplasma che non sedimenta dagli omogenati cellulari, neppure con lĺultracentrifugazione
alle velocitÓ pi¨ elevate.
1.4. Inclusioni citoplasmatiche
Questo termine si riferisce a
strutture, granuli e materiali diversi prodotti dal metabolismo cellulare. Le
inclusioni non fanno parte del gruppo degli organuli, che sono componenti dellĺapparato
cellulare preposti alla sintesi e alla degradazione. Si tratta invece il pi¨
delle volte di sostanze
di riserva sotto forma di granuli, come il glicogeno, di vacuoli
come le inclusioni lipidiche. Le inclusioni comprendono anche i pigmenti, risultato dellĺaccumulo di
prodotti esogeni ed endogeni, degradati o meno, nonchÚ i cristalloidi, formazioni proteiche
che hanno solo lĺapparenza di cristalli.
|
sommario |
avanti |
|
|
|