Vol. 2° - V.1.
Morfologia dei mitocondri
1.1. Morfologia al microscopio ottico
Forma
I mitocondri possono presentarsi
con morfologia variabile: allungati, filamentosi, oppure granulari. La forma
varia con la posizione occupata nella cellula. Cosě, nelle cellule
intestinali hanno una disposizione bipolare: al polo apicale sono filamentosi,
al polo basale si presentano granulari. La pressione osmotica e il pH ne
influenzano la forma: a pH acido tendono ad assumere struttura vescicolare.
Dimensioni
Anche le dimensioni dei
mitocondri possono variare. Nella maggior parte delle cellule si presentano
come bastoncini del diametro di 0,2÷1 μm
e possono raggiungere una lunghezza massima di 10 μm.
Tuttavia la forma dipende dall’attivitŕ cellulare, in quanto i mitocondri
si modificano durante il ciclo secretorio, quando possono avere uno spessore
maggiore, con uno diametro di 0,2 ¸
2 μm
al massimo.
Distribuzione
La ripartizione dei mitocondri
nel citoplasma č molto uniforme. Esistono perň delle eccezioni alla regola.
Nei tubuli contorti del rene si dispongono alla base della cellula e al
microscopio elettronico si presentano come bastoncini basali, perpendicolari
alla membrana basale della cellula, in quanto occupano i compartimenti che
delimitano le invaginazioni della membrana cellulare. Talora si distribuiscono
intorno al nucleo oppure alla periferia del citoplasma, in certi casi si
dispongono intorno al centriolo tra i filamenti dell’aster. La
localizzazione dei mitocondri dipende dalla funzione che la cellula sta
svolgendo, essendo prevalenti in quelle aree in cui la cellula necessita di
energia. Nel muscolo si trovano di fronte ai dischi A delle miofibrille, nei
bastoncelli della retina occupano il segmento interno, nella regione basale
della cellula tubulare renale liberano l’energia che la membrana utilizza
per il trasporto dell’acqua e delle altre molecole.
Quantitŕ
Il numero di mitocondri per
cellula č variabile, essendo in relazione al fabbisogno energetico: mentre in
una normale cellula si trovano da 1.000 a 2.000 mitocondri, nell’oocita di
alcune specie animali essi possono salire a 30.000. Si ritiene che nella
cellula epatica i mitocondri costituiscano dal 30 al 35% delle proteine
cellulari totali.
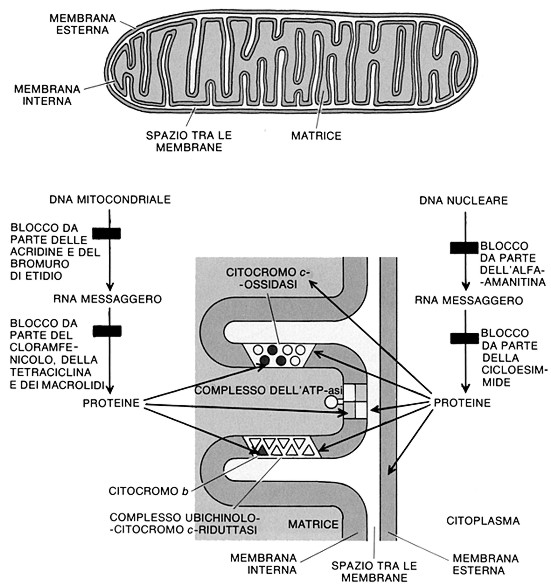
Fig. V. 1 - Struttura schematica di un mitocondrio
1.2. I movimenti dei mitocondri
Nella cellula a riposo
la
microcinematografia accelerata in contrasto di fase secondo Frédéricq e
Chčvremont (1951) mette in evidenza mitocondri animati da movimenti
incessanti, con spostamenti dovuti alle correnti citoplasmatiche di
diffusione, e rapide modificazioni strutturali. I mitocondri si incurvano, si
rimodellano, subiscono un movimento oscillatorio, si attorcigliano, si
strozzano, si frammentano in bastoncelli piů corti oppure si fondono in
bastoncelli piů lunghi. Le correnti citoplasmatiche non intervengono in
queste modificazioni strutturali. Siccome il mitocondrio non č dotato di
contrattilitŕ, le modificazioni della forma sono dovute verosimilmente a
cambiamenti della loro superficie. Gli scambi metabolici fra mitocondri e
citoplasma sarebbero responsabili di reazioni chimiche capaci di determinarne
i movimenti.
Durante la mitosi
il
comportamento dei mitocondri si modifica: i movimenti rallentano e gli
organuli vanno a distribuirsi all’esterno dei complessi cromosomici e
fusali. Durante la citodieresi si ripartiscono nelle cellule figlie in
quantitŕ pressapoco uguale.
1.3. Ultrastruttura dei mitocondri
La costanza strutturale dei
mitocondri, che č indipendente dal tipo di cellula eucariotica, č un dato
decisamente importante.
1.3.a. La parete mitocondriale
Due membrane, interna ed
esterna, separate tra loro da uno spazio, delimitano la camera interna,
fungendo cosě da guscio. Dal punto
di vista strettamente strutturale queste membrane sono elastiche, flessibili e
stabili. La cinematografia in contrasto di fase evidenzia movimenti molto ampi
e deformazioni notevoli a carico dei mitocondri di fibroblasti in coltura.
Queste modificazioni della forma dipendono dalla composizione e dalla
disposizione delle molecole della parete. La costituzione biochimica in lipidi
delle membrane č stata studiata da André e collaboratori.
Procedendo dall’esterno,
troviamo in successione:
ü
membrana
esterna, a struttura trilaminare, formata da proteine intrinseche
globulari (3.500-4.000 per mm˛);
tali proteine rappresentano il 60% della membrana esterna e sono localizzate
nello spessore dello strato lipidico, che a sua volta rappresenta il 40%. Le
proteine si dispongono in modo da delimitare dei pori atti a facilitare il
passaggio bidirezionale di metaboliti con peso molecolare inferiore a 10.000
Da. Una tale permeabilitŕ č confermata dal fatto che la composizione
biochimica della camera esterna č simile a quella del citoplasma. La membrana
esterna contiene numerosi enzimi: alcune transferasi, alcune chinasi, ATP
acetil-CoA sintetasi, citocromo b, NADH, citocromo b reduttasi.
ü membrana interna, la cui organizzazione molecolare č completamente diversa da quella esterna. Non solo ha una particolare ricchezza in proteine (80%) e una relativa povertŕ in lipidi (20%), ma possiede una sua organizzazione molecolare. La membrana interna contiene proteine intrinseche e particelle elementari extramembranarie (messe in evidenza con colorazioni negative) collegate a proteine intrinseche per mezzo di un peduncolo. Contiene gli enzimi responsabili delle reazioni di ossidazione che liberano l’energia necessaria alla fosforilazione ossidativa, cioč alla formazione di adenosin trifosfato o ATP a partire da ADP (adenosin difosfato).
ü
camera interna, che rappresenta la
matrice mitocondriale.
1.3.b. Le creste mitocondriali
Si tratta di invaginazioni della
membrana interna a forma di tubuli o di sacculi che penetrano in profonditŕ
nella matrice mitocondriale. Non si conosce molto bene come tali creste siano
adese né la loro morfologia, anche se vengono rappresentate nelle seguenti
fogge:
§ sotto
forma di bottiglie piatte
oppure di fiaschi,
il cui collo, o peduncolo, sarebbe piů stretto della cresta stessa;
§ sotto
forma di sacculi
le cui pareti, senza strozzature, si continuano con la membrana interna.
Contrariamente al caso precedente, il compartimento interno della cresta
comunica liberamente con la camera esterna, e non attraverso uno stretto
orifizio.
§ sotto
forma di tubuli.
1.3.c. La camera interna
Č lo spazio della matrice
mitocondriale delimitato dalla membrana interna, compreso fra le creste
mitocondriali. La matrice č finemente granulare e la sua densitŕ varia in
funzione dello stato funzionale. Vi si trovano costantemente:
o
molecole di DNA
o
mitoribosomi, o mtRNA, visibili dopo colorazioni negative
o
granulazioni dense, irregolari, con diametro di 50 nm
(accumulo di cationi).
La camera interna contiene tutti gli enzimi coinvolti nel
ciclo di Krebs e quelli della biosintesi degli acidi grassi.
|
sommario |
avanti |
|
|
|