Vol. 2░ - XXVI.1.3.
Il sebo-cheratinocita del pollo
╚ stato stabilito da Freinkel (1972)
e
da Wrench (1980) che lĺepidermide del pollo Ŕ un organo deputato
alla sintesi lipidica. Lavker (1975) ha dimostrato che lĺepidermide
del pollo sintetizza in prevalenza trietilgliceroli e alcuni fosfolipidi.
Le cellule epidermiche del pollo differiscono da quelle
dei mammiferi per il fatto di possedere un apparato di Golgi pi¨ sviluppato -
caratteristica condivisa con la ghiandola dellĺuropigio![]() , con le ghiandole
sebacee dei mammiferi e con le cellule del tessuto grasso, o adipociti - e
questo particolare sviluppo dellĺapparato di Golgi sembra proprio in
relazione allĺattivitÓ di sintesi lipidica.
, con le ghiandole
sebacee dei mammiferi e con le cellule del tessuto grasso, o adipociti - e
questo particolare sviluppo dellĺapparato di Golgi sembra proprio in
relazione allĺattivitÓ di sintesi lipidica.
Le cellule dello strato basale dellĺepidermide presentano ben evidente sia il reticolo endoplasmatico liscio che granulare (REL e REG), ma una volta che esse diventano cellule dello strato spinoso solo il REL rimane evidente. Tutto ci˛ Ŕ ben differente da quanto si pu˛ riscontrare nelle cellule epidermiche dei mammiferi, che hanno un reticolo endoplasmatico scarsamente sviluppato.
Nel caso delle
cellule epidermiche del pollo, il RE sembra essere coinvolto nella lipogenesi:
piccole gocce di grassi appaiono dapprima attorno ai nuclei delle cellule
basali, per poi aumentare di volume e confluire a formare delle strutture
globulari di maggiori dimensioni quando le cellule si spostano negli stati
intermedi. La lipogenesi prende lĺavvio giÓ negli ultimi stadi di sviluppo
embrionale.
|
|
Fig. XXVI. 6. ľ Sequenza delle cellule dellĺepidermide di pollo B cellula basale S cellula dello strato spinoso Ti cellula dello strato transizionale inferiore Ts cellula dello strato transizionale superiore C cellule dello strato corneo L gocciole lipidiche intracellulari e intercellulari diffuse sino alla superficie cutanea. |
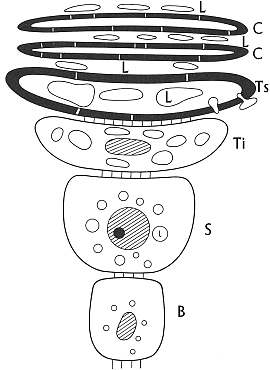
I precursori della cheratina vengono deposti nella parte periferica del citoplasma degli strati epidermici di transizione, e nella parte centrale della cellula di cheratina non se ne forma a causa della presenza di grandi gocciole lipidiche in elevata quantitÓ. Alla periferia della cellula esistono delle aree non cheratinizzate, dovute forse alla diffusione di lipidi in questi punti prima che sia avvenuta la cheratinizzazione.
Attraverso queste lacune, non protette dallĺazione
disintegratrice degli enzimi lisosomiali idrolitici, i grassi riescono a
diffondere nello spazio intercellulare, per cui le cellule cornee contengono
una quantitÓ di grassi inferiore alle cellule degli strati sottostanti.
Le cellule corneificate si trasformano comunque in squame
grassose, per cui lo strato corneo Ŕ come se fosse un sandwich di lipidi,
composto dalle squame al posto delle fette di pane, le quali racchiudono dei
grassi: un panino imbottito di grassi. Ne risulta una cute ricoperta di un
manto costituito da un film lipidico che nel pollo equivale alla secrezione
sebacea dei mammiferi. I lipidi della cute vengono cosý riversati sul
piumaggio.
Lucas & Stettenheim (1972) hanno
potuto dimostrare che nel pollo lĺentitÓ della sintesi lipidica varia da unĺarea
allĺaltra, essendo pi¨ spiccata nelle aree glabre e minima in quelle
impiumate. Ci˛ nonostante, qualsiasi distretto cutaneo mostra unĺattivitÓ
lipidosintetica, e alcuni grassi sono presenti nella guaina del follicolo
della piuma, come pure sulle piume. Le regioni maggiormente attive nella
produzione di grassi sono le seguenti: lĺangolo dĺapertura del becco, la
superficie plantare delle zampe, la cresta e i bargigli, mentre negli apterili
del petto la lipogenesi Ŕ moderata.
Dal punto di vista funzionale lĺepidermide somiglia
molto da vicino allĺuropigio e alla ghiandola sebacea dei mammiferi. Lĺepidermide
dei polli pu˛ essere definita una ghiandola
sebacea olocrina, non discostandosi in questo dalle ghiandole presenti
nellĺuropigio, per cui Ŕ stato coniato il termine di sebo-cheratinocita (Wrench,
1980).
|
sommario |
avanti |
|
|
|