Vol. 2° - XXVII.3.
IL MELANOCITA
Nonostante tutti i melanociti
dei mammiferi condividano la capacitŕ di produrre melanosomi, non tutti i
melanociti si comportano in questo modo, in quanto forma e funzione dipendono in larga misura dalla loro localizzazione.
I melanociti piů numerosi e piů caratteristici sono quelli dell’epidermide,
con i loro dendriti che si insinuano tra i cheratinociti circostanti dentro ai
quali riversano i melanosomi immergendovi le loro estremitŕ. Si tratta di un
processo citocrino non condiviso da tutti i melanociti, e attualmente il punto
di vista č un po’ differente, in quanto si ritiene che i cheratinociti
siano dotati di un’elevata attivitŕ fagocitaria nei confronti dell’estremitŕ
dei dendriti dove sono contenuti i melanosomi.
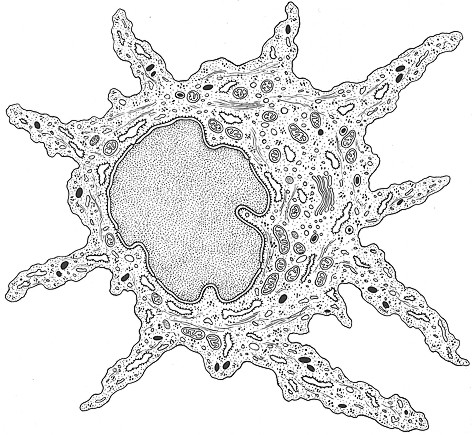
Fig. XXVII. 2. – Ultrastruttura di un melanocita. A causa dei prolungamenti citoplasmatici con aspetto ramificato, queste cellule vengono anche dette cellule dendritiche. Nel contesto del citoplasma sono chiaramente rappresentati i melanosomi.
Il melanocita č una cellula a nucleo piccolo provvista di
prolungamenti dendritici che si insinuano tra le cellule e si anastomizzano
tra loro costituendo una specie di rete. Con i comuni metodi istologici il
citoplasma appare di solito chiaro, ma con appropriati metodi all’argento,
all’oro, alla diidrossifenilalanina, risulta carico di una miriade di
granuli di color bruno piů o meno intenso che occupano anche i dendriti.
Il melanocita č assai facilmente individuabile al microscopio elettronico soprattutto per la mancanza di tonofibrille e di desmosomi. Questo fatto spiega la sua libertŕ e la sua attitudine a insinuarsi tra le cellule epidermiche, le quali, anche quando accolgono i dendriti, rimangono pur sempre saldate tra loro per mezzo di desmosomi che restano invariati.
La ricchezza
dell’apparato mitocondriale e la complessitŕ
del reticolo endoplasmatico suggeriscono che il melanocita deve essere un
elemento dotato di attivitŕ di sintesi vivace. La melanina fabbricata dai
melanociti non solo viene dispersa nell’epidermide o fagocitata dai
cheratinociti della piuma: essa viene inglobata anche da grossi elementi
dermici designati melanofori o cromatofori.
Dopo numerose controversie si č ormai accertato che di
solito la tirosinasi si trova nei melanociti in uno stato di inibizione e che
sono necessarie particolari condizioni per attivarla: quando ciň si verifica,
la tirosinasi attivata provoca la conversione della tirosina in DOPA, inizio
della sintesi melanica.
Una dispersione
dei granuli in tutto il melanocita, anche se la quantitŕ resta
invariata, porta a un’intensificazione del colore e quindi a un
imbrunimento; al contrario, l’aggregarsi
dei granuli nella zona centrale della cellula, in prossimitŕ del
nucleo, rende l’elemento assai piů chiaro e di conseguenza la cute appare
ipo o depigmentata. Questi spostamenti e questo rimaneggiamento dei granuli
possono verificarsi in breve tempo, persino in pochi minuti, e sono sotto il
controllo di sostanze elaborate dall’ipofisi e dalle terminazioni nervose,
forse anche dalla pineale.
I precursori del pigmento sono incolori e acquistano la
tipica tonalitŕ bruna con l’ossidazione. Anche la cute del cadavere puň imbrunire dopo
irradiazione,
fenomeno di Meirowsky,
ma č degno di nota e alquanto singolare il fatto che un’ulteriore
ossidazione tende a decolorare la melanina: ciň sta alla base della
decolorazione dei capelli con acqua ossigenata e spiega le parziali acromie
che si possono notare in capigliature scure dopo prolungata esposizione a luce
solare intensa.
Nei vertebrati omeotermi il melanocita č l’unica cellula in grado di produrre pigmento
melanico. Nel 1948 il canadese Pierre Masson affermň, in modo
profetico, che il melanocita č una cellula
ghiandolare e che il suo prodotto di sintesi viene riversato in un’altra
cellula rappresentata dai cheratinociti e dai macrofagi del derma. Questo
processo di secrezione non č né endocrino né esocrino, bensě citocrino. Sia nei melanociti che nelle
cellule entro le quali viene riversato, il prodotto di secrezione va incontro
a trasformazioni caratteristiche.
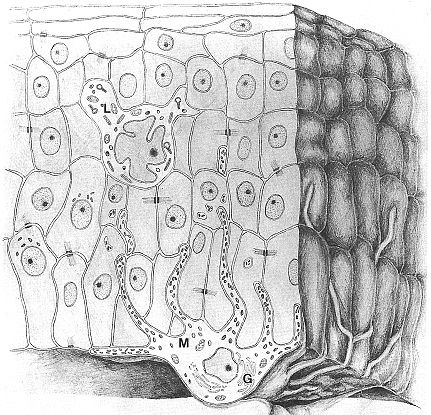
Fig. XXVII. 3. – Melanocita circondato dai cheratinociti satelliti. Il melanocita – M – manda i suoi dendriti tra le cellule dell’epidermide e il suo apparato di Golgi – G – č notevolmente sviluppato in rapporto alla sintesi di melanina. Nel contesto dell’epidermide č rappresentata anche una cellula di Langerhans – L – il cui citoplasma contiene granuli foggiati a racchetta da tennis.
Il sistema melanocitario dei vertebrati superiori, come
gli uccelli e i mammiferi, consiste di due tipi distinti dal punto di vista
anatomico e, almeno sotto certi profili, anche fisiologico:
melanociti dello strato basale dell’epidermide
melanociti dei bulbi piliferi e del follicolo della piuma.
Nell’uomo il numero di melanociti presenti nelle differenti aree del corpo varia in modo spiccato, con una media totale compresa intorno a 1.000÷1.500 mm˛.
Le aree piů densamente popolate sono quelle esposte alla luce solare e certe aree che non lo sono affatto, o che lo sono sempre meno per motivi etologici, come l’area genitale.
Le
differenze nell’ambito di una specie non sono dovute a fattori anatomici,
bensě funzionali, dal momento che il numero dei melanociti č identico senza alcun
rapporto col colore della pelle ed č geneticamente
determinato.
|
sommario |
avanti |
|
|
|