Vol. 2░ - XXVII.5.2.
Ultrastruttura dei granuli di melanina
La struttura dei melanosomi pu‗
essere studiata solo al microscopio elettronico a trasmissione, che ha un
potere dÆingrandimento fino a 1 milione di volte. La melanina contenuta nel
melanosoma Þ presente sotto forma di particelle che non hanno struttura
uniforme, in quanto assumono una morfologia che corrisponde ai vari stadi del
loro sviluppo, e quindi hanno un aspetto che Þ variabile in rapporto al
momento in cui vengono osservate.
5.2.a. Premelanosoma
Il premelanosoma, dotato di
struttura granulare, funge da intermediario. Durante il suo sviluppo il
premelanosoma va incontro a diverse trasformazioni: le forme pi¨ precoci
contengono filamenti spiralizzati relativamente poco elettrondensi;
successivamente il premelanosoma mostra un tenue incremento della densitÓ
elettronica e un profondo rimodellamento della sua struttura fibrillare.
Vediamo di analizzare nei dettagli queste due tappe importanti della sintesi
melanica.
Premelanosoma
fibrillare. Questa struttura precoce contiene fibrille
spiralizzate disposte in modo relativamente lasso allÆinterno di una
vescicola la cui parete Þ liscia. I filamenti stessi delineano il profilo
ovale caratteristico del futuro melanosoma maturo. Il passo delle eliche Þ
pari a circa 9¸10 nm. I filamenti possono rimanere separati, ma in certi
premelanosomi si pu‗ anche osservare una tendenza di due o pi¨ filamenti a
unirsi per dar luogo a un filamento di diametro maggiore. La periodicitÓ
assiale delle fibre spiralizzate oscilla intorno a 6,5¸7,6 nm.
Questo tipo di granulo sembra precedere le altre forme, e
nel suo contesto non mostra ancora la deposizione di materiale elettrondenso.
Si pu‗ affermare con una certa sicurezza che questa struttura fibrillare
definisce la forma del melanosoma e include siti di tirosinasi ancora
inattivi.
Premelanosoma
striato. In uno stadio successivo in certe aree del
premelanosoma si osserva la comparsa di una fine striatura trasversale, e
precisamente lÓ dove le fibre spiralizzate sembrano fondersi in filamenti
pi¨ spessi o in un corpo centrale condensato. LÆaumento della densitÓ
elettronica osservabile in questa fase di sviluppo pu‗ corrispondere a una
concomitante deposizione di melanina elettrondensa. ╚ ragionevole supporre
che la condensazione delle fibrille e lÆattivazione della tirosinasi siano
due processi con alcuni fattori in comune e che essi inizino
contemporaneamente. Non si pu‗ tralasciare di accennare a unÆaltra
possibile spiegazione dellÆincremento della densitÓ elettronica. Infatti,
la stessa tirosinasi forma una struttura cristallina e causa la deposizione di
materiale elettrondenso sulla sua superficie. Anche molecole di metalli
possono causare un aumento della densitÓ, ed Þ noto che lo zinco si accumula
nei granuli di melanina in concentrazione maggiore rispetto alla maggioranza
degli altri componenti della cellula.
La formazione di melanina non ha luogo fintanto che lÆorientamento
delle fibre nel contesto della matrice non sia completato, e la sintesi di
pigmento prosegue sin quando i dettagli della matrice non vengono
completamente persi a causa dalla melanina elettrondensa.
5.2.b. Melanosoma
Sezionando un melanosoma si
possono distinguere due regioni:
Core
centrale. In sezioni ultrasottili di materiale fissato con
tetrossido di osmio si pu‗ mettere in evidenza una striatura longitudinale.
Corticale
osmiofila. La scomparsa della zona corticale compatta e
osmiofila del melanosoma, dopo trattamento sbiancante con permanganato, mette in evidenza la natura melanica
della sostanza osmiofila che compone la corteccia. In condizioni favorevoli,
in seno alla corteccia si pu‗ distinguere una fine struttura che pu‗ essere
correlata alla struttura del polimero
di melanina, che consiste in piccoli granuli disposti in modo
regolare, pi¨ o meno aggregati in unitÓ del diametro di
30¸50 nm. Drochmans ha proposto per
queste formazioni granulari il termine di particelle di melanina.
La stessa sostanza cosý strutturata Þ spesso osservabile
a carico dei melanosomi durante la loro degradazione che si svolge nei vacuoli
di fagocitosi dei melanofagi del derma, dove le particelle di melanina vengono
disgregate in particelle pi¨ piccole. QuestÆosservazione circa lo
spostamento dei melanosomi in seno ai macrofagi sembra essere in
contraddizione con la resistenza della melanina agli agenti chimici in vitro e
con la mancanza di enzimi capaci di depolimerizzarla in vivo. Si pu‗ tuttavia
supporre che polimeri dotati di peso molecolare relativamente basso, intorno a
106, corrispondenti alle fini strutture osmiofile, vengano
assemblati in particelle di dimensioni ridotte, successivamente disposte alla
periferia del melanosoma. Il rilascio di particelle melaniche da parte del
melanosoma durante la degradazione non implica una depolimerizzazione ma solo
una manifestazione di differenti livelli dellÆorganizzazione
macromolecolare.
In sintesi, possiamo affermare che la formazione di un melanosoma
si svolge nel modo seguente:
À
formazione di una matrice proteica composta da filamenti
spiralizzati
À
successiva deposizione di materiale elettrondenso che dÓ
luogo a una struttura fine, verosimilmente dovuta a melanina
À
estensione e limitazione della deposizione di melanina.
Queste osservazioni suggeriscono lÆesistenza nei
melanosomi di differenti
gradi di organizzazione del polimero di melanina:
º
un primo grado di organizzazione macromolecolare Þ
visibile al microscopio elettronico sotto forma di un assemblamento di fini
granuli disposti secondo una struttura regolare
º
il secondo livello di organizzazione consiste nella
formazione di particelle del diametro di 30¸50 nm
º
queste particelle si dispongono a conchiglia intorno allÆunitÓ
elementare, il melanosoma
º
finalmente, microscopicamente parlando, i diversi
melanosomi si congregano a formare un complesso melanosomico.
Essendo talora difficile distinguere tra melanosomi e
premelanosomi, si potrebbe adottare la proposta di Fitzpatrick (1966):
o
Premelanosoma:
qualsiasi formazione che appartiene al processo di maturazione del melanosoma,
caratterizzato da attivitÓ tirosinasica dopo che la sintesi melanica ha preso
lÆavvio; Þ dotato di densitÓ elettronica variabile.
o
Melanosoma:
organulo distinto, contenente melanina, in cui il processo di sintesi melanica
Þ completo; lÆattivitÓ tirosinasica non Þ dimostrabile nel suo contesto e
la densitÓ elettronica Þ pi¨ o meno uniforme.
o
Complessi
melanosomici: si tratta di molteplici melanosomi incastonati su una
matrice di supporto, come accade nei macrofagi e nelle cellule malpighiane dei
mammiferi.
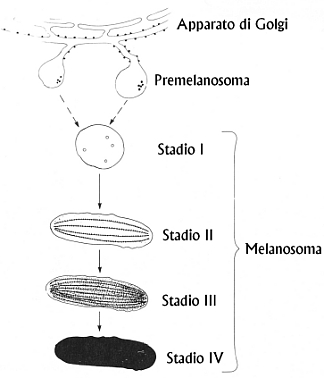
Fig. XXVII. 5. û
Formazione e maturazione del melanosoma
secondo Montagna
Secondo una tassonomia pi¨ recente suggerita da Montagna,
i vari stadi di formazione e maturazione del melanosoma possono essere
schematizzati nel modo seguente:
º
Melanosoma
stadio 1. Appena formati, i premelanosomi sono rotondeggianti
oppure ovalari, e si tratta di vescicole delimitate da una membrana contenente
tirosinasi e filamenti con una periodicitÓ di 9¸10 nm.
º
Melanosoma stadio 2. Si presentano come corpuscoli ovalari anchÆessi
contenenti filamenti con periodicitÓ di 9¸10 nm
º
Melanosoma stadio 3. La melanina sintetizzata al loro interno offusca i
dettagli strutturali; la loro forma Þ sempre ovale.
º
Melanosoma
stadio 4. Si tratta sempre di melanosomi ovalari contenenti
melanina in quantitÓ tale che tutti i dettagli strutturali vengono
completamente mascherati.
Le dimensioni e lo stadio fino al quale i melanosomi riescono a
svilupparsi dipendono dalla costituzione genetica individuale, in
quanto gli individui di pelle pi¨ scura producono melanosomi pi¨ grandi e
pi¨ maturi rispetto ai soggetti di pelle chiara. Come abbiamo giÓ avuto modo
di precisare, nei Bianchi i melanosomi sono piccoli e gli aggregati che essi
formano sono contornati da una membrana.
Numerosi autori hanno spiegato la limitazione della
crescita del melanosoma come dovuta alla deposizione di pigmento e conseguente
seppellimento dellÆenzima in seno
alla struttura finale. Ipotesi interessante, dovuta verosimilmente al fatto
che la formazione di un certo numero di particelle elementari di una
determinata dimensione sia giÓ di per sÚ il fattore limitante.
A scopo riassuntivo, riportiamo
la classificazione dovuta a Fitzpatrick (1965) relativa alle cellule dei
vertebrati contenenti melanina:
À
melanoblasto:
Þ la cellula che, durante tutti gli stadi del ciclo vitale, funge da
precursore dei melanociti e dei melanofori; esso diventa melanocita con la
formazione di melanina o di premelanosomi privi di melanina
À
melanocita:
cellula deputata alla sintesi di un organulo specializzato contenente
melanina, il melanosoma
À
melanoforo:
si tratta di un tipo di melanocita che partecipa, insieme ad altri
cromatofori, ai rapidi mutamenti cromatici degli animali a sangue freddo,
attraverso lÆaggregazione o la dispersione dei melanosomi
À
melanocita
albino: come altri melanociti, caratterizzati da un prodotto
finale costituito da premelanosomi solo parzialmente melanizzati, appartiene
ai melanociti, e i suoi premelanosomi sono totalmente privi di melanina.
|
sommario |
avanti |
|
|
|