Lessico
Pinguino

Pinguino saltatore - Eudyptes chrysocome
![]()
Pinguino deriva dal francese pingouin, che forse risale al bretone penngwenn, testa bianca. Denominazione comunemente attribuita a tutti gli Uccelli Sfenisciformi della famiglia Sfeniscidi. A quasi tutte le specie, tuttavia, vanno attribuite altre denominazioni piů pertinenti (pigoscelide, megadipte, eudipte, eudiptola, sfenisco).

Pinguino
reale - Aptenodytes patagonicus
Picture:
© Samuel Blanc / www.sblanc.com
Il pinguino reale (Aptenodytes patagonicus) č una delle specie piů grandi della famiglia: raggiungendo la lunghezza di 95 cm e una lunghezza degli arti anteriori natatori di oltre 30 cm. Ha becco lungo, sottile e lievemente ricurvo verso il basso, zampe ricoperte di piume sino all'attacco delle dita, che sono munite di unghie ben sviluppate. Grazie a Giovanni Boano possiamo sapere che il dito posteriore, o 1° dito, č vestigiale, cioč rudimentale (Del Hoyo et al., 1992 - Handbook of the birds of the World, Vol. I) e che le altre tre, ben sviluppate, sono unite da membrana interdigitale.
La coda, ridottissima, č formata da penne trasformate in strutture bastoncellari. Il piumaggio č nero sul capo, sulla nuca e sulla gola, mentre in corrispondenza della regione delle orecchie, sul collo e in parte del petto č di un bel colore giallo. Per il resto č grigio nelle regioni superiori del corpo, bianco in quelle inferiori. Il becco č nero e rosso, le zampe brune. Il pinguino reale č diffuso in Patagonia, Georgia del Sud, isole Falkland e negli altri arcipelaghi prossimi al margine occidentale dell'Antartide, dove vive in enormi branchi. Dotato di spessi depositi adiposi sottocutanei, puň galleggiare senza quasi compiere movimenti e nuotare velocemente compiendo frequenti immersioni alla ricerca di pesci e di altri organismi marini di cui si nutre.

Pinguino
imperatore - Aptenodytes forsteri
Picture:
© Samuel Blanc / www.sblanc.com
Simile al precedente ma di taglia piů grande č il pinguino imperatore (Aptenodytes forsteri![]() ), che
raggiunge la lunghezza di 115 cm e abita esclusivamente le coste antartiche.
), che
raggiunge la lunghezza di 115 cm e abita esclusivamente le coste antartiche.
Il pinguino imperatore si riproduce nel pieno dell'inverno antartico, raggiungendo verso la fine di marzo i luoghi di nidificazione, che si trovano a parecchie decine di chilometri nell'entroterra, spostandosi in grandi stuoli. Č possibile che alcune coppie si riformino di anno in anno (i due partners possono riconoscere il rispettivo richiamo fra la moltitudine), ma il meccanismo di formazione delle coppie non č completamente conosciuto. Come in altri pinguini, l'uovo, che in questa specie č singolo e viene deposto fra maggio e giugno, č preso in consegna dal maschio, che lo sostiene sul dorso dei piedi e lo ricopre con una piega della pelle ventrale, isolandolo dal freddo intensissimo dell'ambiente esterno. Le femmine della colonia si riuniscono allora in gruppi e si dirigono verso il mare e, a seconda della regolaritŕ del ghiaccio, camminano erette o scivolano sul ventre. Giunte al mare, spesso dopo parecchi giorni di cammino, esse vi trascorrono diversi giorni, nutrendosi soprattutto di crostacei, cefalopodi e pesci e ricostituendo le spesse riserve di grasso sottocutaneo diminuite nel corso della marcia per e dai luoghi di nidificazione e dalla riproduzione. Dopodiché tornano presso i loro compagni.
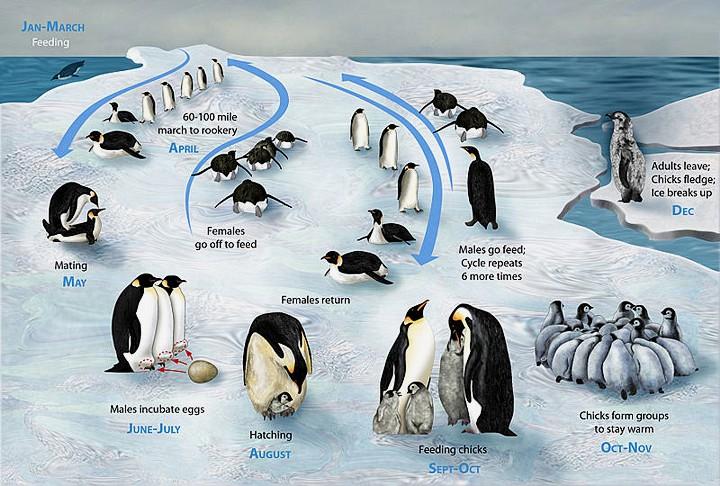
I maschi che incubano le uova non si nutrono e vivono a spese delle loro riserve di grasso; quando le condizioni climatiche lo consentono essi possono muoversi di qua e di lŕ senza mai abbandonare il loro uovo; ma nei periodi di maltempo, con temperature di parecchie decine di gradi sotto zero, si ammassano strettamente fra loro esponendo alle intemperie una superficie assai ridotta: i pinguini che si trovano in posizione piů periferica rivolgono al vento il dorso e senza sosta premono per entrare nel mezzo del gruppo, avvicendandosi continuamente nelle posizioni piů esposte e in quelle piů riparate. Pare che nei periodi in cui la temperatura dell'ambiente esterno si approssima ai limiti minimi, il metabolismo basale dei pinguini, e quindi la loro temperatura interna, si abbassino, adattamento che riduce sia il consumo del grasso che la perdita di calore.
Le uova si schiudono in poco piů di due mesi, appena dopo il rientro delle femmine o, se l'assenza di queste si prolunga, poco prima; in questo caso pare che alcuni padri siano in grado di nutrire i loro piccoli con un po' di cibo indigerito che ancora conservano nel gozzo; ma in genere i piccoli escono dalle uova fra i piedi delle madri, a cui i maschi cedono la cura delle uova, che li nutrono con cibo fresco e abbondante.
Al ritorno delle femmine, i maschi, notevolmente dimagriti (in casi estremi arrivano a perdere fino alla metŕ del loro peso iniziale), intraprendono a loro volta il viaggio verso il mare per mangiare e per raccogliere cibo da portare ai piccoli, alternandosi con le madri nella protezione della prole e nei viaggi di approvvigionamento. I piccoli, che nei primi giorni di vita hanno una termoregolazione imperfetta, vengono protetti fra i piedi e il ventre dei genitori fino alla fine dell'inverno. All'etŕ di due mesi i giovani, ancora ricoperti di piumino grigio, si uniscono ad altri giovani, ma continuano a essere alimentati dai genitori, ai cui richiami accorrono quando essi tornano dal mare col gozzo carico di cibo. A cinque mesi di etŕ i giovani incominciano a formare le penne definitive, la cui copertura si completa in poco piů di un mese; a questa etŕ essi sono pronti a entrare in mare, dove imparano la coordinazione del nuoto e dell'immersione, compiono le prime esperienze di pesca e imparano a sfuggire ai predatori.
Gli adulti, dopo un periodo di nutrizione intensa, si riportano a terra per compiere la muta delle penne, che avviene contemporaneamente in tutto il corpo; in seguito tornano ancora al mare, che in questo periodo, corrispondente alla fine dell'estate e all'autunno, č particolarmente ricco di alimenti, e di nuovo accumulano grandi quantitŕ di grasso col quale sopravviveranno nell'inverno e nella stagione riproduttiva successivi.
Aptenodytes forsteri
Pinguino imperatore

Uovo di pinguino imperatore
Classe: Aves
Ordine: Sphenisciformes
Famiglia: Spheniscidae
Genere: Aptenodytes
Specie: forsteri, Gray,
1844
Il pinguino imperatore - Aptenodytes forsteri (descritto da George Robert Gray nel 1844, dopo la sua scoperta nei mari antartici), č il piů grande degli uccelli appartenenti alla famiglia dei pinguini (Spheniscidae), insieme al pinguino reale (Aptenodytes patagonicus) appartiene al genere dei pinguini di grandi dimensioni (Aptenodytes), nessuno capace di volare ma tutti ottimi nuotatori. Lo zoologo inglese George Robert Gray nel 1844, dopo averlo scoperto nei mari antartici, grazie al greco antico lo denominň aptenodytes (aptën, genitivo aptęnos = senza ali e dýtës = tuffatore), mentre forsteri ricorda il naturalista tedesco Johann Reinhold Forster che aveva accompagnato il capitano James Cook durante il suo secondo viaggio nel Pacifico (1772-75) quando aveva battezzato 5 specie di pinguini.
Il pinguino imperatore č alto in media 115 cm, per un peso compreso fra i 22 e i 47 kg a seconda della fase del ciclo riproduttivo in cui si trova: i pinguini imperatore tendono infatti a perdere peso durante l'incubazione dell'uovo e durante la cura dei piccoli. Č la specie di pinguino piů grande esistente. La testa, il collo, il dorso e la parte dorsale delle ali sono nere, con l'approssimarsi del periodo della muta il colore tende a sbiadire verso il marrone. La parte ventrale e il lato interno delle ali sono di colore bianco. Nella zona auricolare presenta delle macchie arancione tondeggianti che sfumano verso il bianco a mano a mano che si uniscono alla parte ventrale. Le zampe sono nere o comunque scure. Il becco č relativamente lungo e presenta la parte superiore di colore nero mentre quella inferiore č arancione, rosa o color lillŕ.
Non vi sono differenze di dimensione e di colore fra maschi e femmine, mentre gli individui piů giovani si distinguono per il becco piů scuro e una colorazione piů chiara delle macchie auricolari e per il collo bianco. I piccoli sono coperti di piume grigio-argentee con testa nera e zona oculare e auricolare bianca.
Il pinguino imperatore č la specie di pinguini che vive piů a sud, i circa 400.000 esemplari vivono sui ghiacci nei mari antartici in latitudini comprese fra i 66° e 78° S. Si riproduce sui ghiacci che circondano le coste fino a 18 km dalla costa, vi sono solo due colonie note che si riproducono a terra. Le colonie in riproduzione tendono a scegliere le zone ghiacciate che presentano dei rilievi in grado di proteggerli dal vento. Per nutrirsi, si spinge in mare aperto.
Il pinguino imperatore č un ottimo nuotatore, raggiunge in acqua velocitŕ di 3,4 metri al secondo (12,2 km/h) e riesce a immergersi fino a 400/450 m di profonditŕ rimanendo sott'acqua fino a 22 minuti. Le immersioni medie dei pinguini sono effettuate fra i 150 e 200 m e durano tra i 3 e i 6 minuti. Non č noto quale sia il metodo di rilevamento delle prede alle alte profonditŕ in assenza di luce. La parte piů consistente dell'alimentazione del pinguino imperatore č costituita da crostacei amfipodi e da krill, si nutrono inoltre di pesci (Gymnodraco acuticeps, Pleuragramma antarcticum, diversi Nototheniidae) e di cefalopodi (Psychroteuthis glacialis, Alluroteuthis antarcticus e Kondakovia longimana).
Tra marzo e aprile i pinguini imperatore si riuniscono in vaste colonie, nel corso della stagione riproduttiva sono monogami. Benché gli individui formino una coppia con un individuo diverso ogni anno, uno studio ha rilevato che il 14,6% delle coppie si ricostituisce l'anno successivo e il 4,9% nuovamente per il terzo anno. Le femmine tra maggio e giugno depongono un solo uovo di grosse dimensioni (460/470 grammi) che dopo poco tempo viene passato al maschio per la cova mentre la femmina ritorna ai suoi usuali territori di pesca. L'incubazione avviene nel pieno dell'inverno antartico quando la temperatura puň scendere anche a -60 °C e con venti che possono raggiungere i 200 km/h. Il maschio nasconde l'uovo sopra le sue zampe e lo protegge ricoprendolo con una sorta di tasca ventrale. Durante la cova il maschio digiuna, il suo peso corporeo si riduce in maniera notevole, l'incubazione dura dai 62 ai 67 giorni. Se il piccolo nasce prima del ritorno della madre, il maschio lo nutre con una sorta di secrezione gastrica biancastra, al rientro della madre il pinguino maschio torna a pescare e a nutrirsi, in assenza del padre la madre nutre il piccolo con pesce rigurgitato, il piccolo viene poi nutrito da entrambi i genitori fino all'indipendenza, ovvero intorno ai 4 mesi. I giovani pinguini si aggregano in gruppi detti crčche attendendo l'arrivo dei genitori che portano il cibo. I pinguini nutrono solo il loro piccolo, il riconoscimento avviene tramite una sorta di fischio modulato emesso dal piccolo e udibile a lunga distanza.
Una volta lasciata la colonia, pare che i pinguini si disperdano in mare aperto ma sono rari gli individui trovati piů a nord di 60° di latitudine sud. Sono stati avvistati esemplari in Nuova Zelanda, nelle regioni piů meridionali dell'America del Sud oltre alle isole subantartiche.
La specie non appare in generale in pericolo di estinzione. Le stime del 1995 contano 195.400 coppie (tra i 400 000 e i 450 000 esemplari) e il numero appare stabile anche se vi sono colonie in declino come a Capo Crozier dove il numero č in diminuzione fin dagli anni settanta, o a Punta Geologia (Terra di Adélie), dove tra il 1950 e il 1975 erano presenti 6.000 coppie circa, ridottosi a 2.500 coppie nel 1982. I pinguini imperatori sono disturbati soprattutto dal sorvolo degli elicotteri di passaggio per il rifornimento alle basi scientifiche. Il surriscaldamento globale, inoltre, sta riducendo le aree ghiacciate anche in Antartide.
Penguins (order Sphenisciformes, family Spheniscidae) are a group of aquatic, flightless birds living almost exclusively in the southern hemisphere, especially in Antarctica, where they are most well-known for living. Highly adapted for life in the water, penguins have countershaded dark and white plumage, and their wings have become flippers. Most penguins feed on krill, fish, squid, and other forms of sealife caught while swimming underwater. They spend about half of their life on land and half in the oceans. Although all penguin species are native to the southern hemisphere, they are not found only in cold climates, such as Antarctica. In fact, only a few species of penguin live so far south. Several species are found in the temperate zone, and one species, the Galápagos Penguin, lives near the equator.
The largest living species is the Emperor Penguin (Aptenodytes forsteri![]() ):
adults average about 1.1 m (3 ft 7 in) tall and weigh 35 kg (75 lb) or more.
The smallest penguin species is the Little Blue Penguin (also known as the
Fairy Penguin), which stands around 40 cm tall (16 in) and weighs 1 kg (2.2 lb).
Among extant penguins, larger penguins inhabit colder regions, while smaller
penguins are generally found in temperate or even tropical climates. Some
prehistoric species attained enormous sizes, becoming as tall or as heavy as
an adult human (see below for more). These were not restricted to Antarctic
regions; on the contrary, subantarctic regions harboured high diversity, and
at least one giant penguin occurred in a region not quite 2,000 km south of
the equator, in a climate decidedly warmer than today.
):
adults average about 1.1 m (3 ft 7 in) tall and weigh 35 kg (75 lb) or more.
The smallest penguin species is the Little Blue Penguin (also known as the
Fairy Penguin), which stands around 40 cm tall (16 in) and weighs 1 kg (2.2 lb).
Among extant penguins, larger penguins inhabit colder regions, while smaller
penguins are generally found in temperate or even tropical climates. Some
prehistoric species attained enormous sizes, becoming as tall or as heavy as
an adult human (see below for more). These were not restricted to Antarctic
regions; on the contrary, subantarctic regions harboured high diversity, and
at least one giant penguin occurred in a region not quite 2,000 km south of
the equator, in a climate decidedly warmer than today.
The word Penguin is thought by some to derive from the Welsh words pen (head) and gwyn (white), applied to the Great Auk which had white spots in front of its eyes (although its head was black); or from an island off Newfoundland known as Pengwyn, due to its having a large white rock. (In the latter case, the name may also have come from Breton.) This theory is supported by the fact that penguins look remarkably like Great Auks in general shape. It is also possible that penguin comes from the Latin pinguis, “fat”. This is supported by the fact that the corresponding words in most other languages (e.g., French pingouin, German Pinguin) have i instead of e as the first vowel. However, a Welsh i is often sound-shifted to an e in the English language. Another theory states that the word is an alteration of “pen-wing”, with reference to the rudimentary wings of both Great Auks and penguins, but there is no evidence to support this.
What may be a King Penguin but certainly is a member of the Spheniscidae
appears on a 1599 map at the Strait of Magellan![]() with the caption "Pinguyn".
The map's features are labeled in Latin, such as Fretum Magellanicum ("Strait
of Magellan"). In addition, there is ample evidence that the Latin term
anser magellanicus ("Goose of Magellan" or "Magellanic
Goose") was the usual term for penguins in the scholarly literature of
that time. In a final twist to the story, the term "Magellanic
Goose" (today usually "Magellan Goose") in our time has come to
denote an actual anseriform, namely a Chloephaga sheldgoose.
with the caption "Pinguyn".
The map's features are labeled in Latin, such as Fretum Magellanicum ("Strait
of Magellan"). In addition, there is ample evidence that the Latin term
anser magellanicus ("Goose of Magellan" or "Magellanic
Goose") was the usual term for penguins in the scholarly literature of
that time. In a final twist to the story, the term "Magellanic
Goose" (today usually "Magellan Goose") in our time has come to
denote an actual anseriform, namely a Chloephaga sheldgoose.

Spheniscus magellanicus
Living species and recent extinctions - Emperor Penguins (Aptenodytes forsteri), the largest living species. Adelie Penguin (Pygoscelis adeliae) feeding young. Like its relatives, a neatly bi-coloured species with a head marking. Magellanic Penguins (Spheniscus magellanicus) guarding nest burrow. The closed neck collar denotes this species. Closeup of Southern Rockhopper Penguin (Eudyptes chrysocome).The number of extant penguin species is debated. Depending on which authority is followed, penguin biodiversity varies between 17 and 20 living species, all in the subfamily Spheniscinae. Some sources consider the White-flippered Penguin a separate Eudyptula species, while others treat it as a subspecies of the Little Penguin; the actual situation seems to be more complicated. Similarly, it is still unclear whether the Royal Penguin is merely a color morph of the Macaroni penguin. Also eligible to be a separate species is the Northern population of Rockhopper penguins.

Aptenodytes patagonicus

Updated after Marples (1962), Acosta Hospitaleche (2004), and Ksepka et al. (2006).
Subfamily Spheniscinae – Modern penguins
Aptenodytes – Great penguins
King Penguin, Aptenodytes patagonicus
Emperor Penguin, Aptenodytes forsteri
Pygoscelis – Brush-tailed penguins
Adelie Penguin, Pygoscelis adeliae
Chinstrap Penguin, Pygoscelis antarctica
Gentoo Penguin, Pygoscelis papua
Eudyptula – Little penguins
Little Blue Penguin, Eudyptula minor
Northern Little Penguin, Eudyptula albosignata (provisional)
Spheniscus – Banded penguins
Magellanic Penguin, Spheniscus magellanicus
Humboldt Penguin, Spheniscus humboldti
Galapagos Penguin, Spheniscus mendiculus
African Penguin, Spheniscus demersus
Megadyptes
Yellow-eyed Penguin, Megadyptes antipodes
Waitaha Penguin, Megadyptes waitaha (extinct)
Eudyptes – Crested penguins
Fiordland Penguin, Eudyptes pachyrynchus
Snares Penguin, Eudyptes robustus
Erect-crested Penguin, Eudyptes sclateri
Southern Rockhopper Penguin, Eudyptes chrysocome
Northern Rockhopper Penguin, Eudyptes moseleyi
Royal Penguin, Eudyptes schlegeli (disputed)
Macaroni Penguin, Eudyptes chrysolophus
Chatham Islands Penguin, Eudyptes sp. (extinct)
Some recent sources apply the phylogenetic taxon Spheniscidae to what here is referred to as Spheniscinae. Furthermore, they restrict the phylogenetic taxon Sphenisciformes to flightless taxa, and establish the phylogenetic taxon Pansphenisciformes as equivalent to the Linnean taxon Sphenisciformes, i.e., including any flying basal "proto-penguins" to be discovered eventually. Given that neither the relationships of the penguin subfamilies to each other nor the placement of the penguins in the avian phylogeny is presently resolved, this seems spurious [neutrality disputed] and in any case is confusing; the established Linnean system is thus followed here.
The evolutionary history of penguins is well-researched and represents a showcase of evolutionary biogeography; though as penguin bones of any one species vary much in size and few good specimens are known, the alpha taxonomy of many prehistoric forms still leaves much to be desired. Some seminal articles about penguin prehistory have been published since 2005, the evolution of the living genera can be considered resolved by now. The basal penguins lived around the time of the Cretaceous–Tertiary extinction event somewhere in the general area of (southern) New Zealand and Byrd Land, Antarctica. Due to plate tectonics, these areas were at that time less than 1,500 kilometers (932 mi) apart rather than the 4,000 kilometers (2,485 mi) of today. The most recent common ancestor of penguins and their sister clade can be roughly dated to the Campanian–Maastrichtian boundary, around 70–68 mya. What can be said as certainly as possible in the absence of direct (i.e., fossil) evidence is that by the end of the Cretaceous, the penguin lineage must have been evolutionarily well distinct, though much less so morphologically; it is fairly likely that they were not yet entirely flightless at that time, as flightless birds have generally low resilience to the breakdown of trophic webs which follows the initial phase of mass extinctions because of their below-average dispersal capabilities (see also Flightless Cormorant).
The oldest known fossil penguin species is Waimanu manneringi, which lived in the early Paleocene epoch of New Zealand, or about 62 mya. While they were not as well-adapted to aquatic life as modern penguins, Waimanu were generally loon-like birds but already flightless, with short wings adapted for deep diving. They swam on the surface using mainly their feet, but the wings were – as opposed to most other diving birds, living and extinct – already adapting to underwater locomotion.
Perudyptes from northern Peru was dated to 42 mya. An unnamed fossil from Argentina proves that by the Bartonian (Middle Eocene), some 39–38 mya, primitive penguins had spread to South America and were in the process of expanding into Atlantic waters.
During the Late Eocene and the Early Oligocene (40–30 mya), some lineages of gigantic penguins existed. Nordenskjoeld's Giant Penguin was the tallest, growing nearly 1.80 meters (6 ft) tall. The New Zealand Giant Penguin was probably the heaviest, weighing 80 kg or more. Both were found on New Zealand, the former also in the Antarctic farther eastwards.
Traditionally, most extinct species of penguins, giant or small, had been placed in the paraphyletic subfamily called Palaeeudyptinae. More recently, with new taxa being discovered and placed in the phylogeny if possible, it is becoming accepted that there were at least two major extinct lineages. One or two closely related ones occurred in Patagonia, and at least one other – which is or includes the paleeudyptines as recognized today – occurred on most Antarctic and subantarctic coasts.
But size plasticity seems to have been great at this initial stage of penguin radiation: on Seymour Island, Antarctica, for example, around 10 known species of penguins ranging in size from medium to huge apparently coexisted some 35 mya during the Priabonian (Late Eocene). It is not even known whether the gigantic palaeeudyptines constitute a monophyletic lineage, or whether gigantism was evolved independently in a much restricted Palaeeudyptinae and the Anthropornithinae – whether they were considered valid, or whether there was a wide size range present in the Palaeeudyptinae as delimited as usually done these days (i.e., including Anthropornis nordenskjoeldi). The oldest well-described giant penguin, the 5-foot-tall Icadyptes salasi, actually occurred as far north as northern Peru about 36 mya.
In any case, the gigantic penguins had disappeared by the end of the Paleogene, around 25 mya. Their decline and disappearance coincided with the spread of the Squalodontoidea and other primitive, fish-eating toothed whales, which certainly competed with them for food, and were ultimately more successful. A new lineage, the Paraptenodytes which includes smaller but decidedly stout-legged forms, had already arisen in southernmost South America by that time. The early Neogene saw the emergence of yet another morphotype in the same area, the similarly-sized but more gracile Palaeospheniscinae, as well as the radiation which gave rise to the penguin biodiversity of our time.
Modern penguins constitute two undisputed clades and another two more basal
genera with more ambiguous relationships. The origin of the Spheniscinae
lies probably in the latest Paleogene, and geographically it must have been
much the same as the general area in which the order evolved: the oceans
between the Australia-New Zealand region and the Antarctic. Presumedly
diverging from other penguins around 40 mya, it seems that the Spheniscinae
were for quite some time limited to their ancestral area, as the
well-researched deposits of the Antarctic Peninsula and Patagonia have not
yielded Paleogene fossils of the subfamily. Also, the earliest spheniscine
lineages are those with the most southern distribution.
The genus Aptenodytes appears to be the basal most divergence among
living penguins; they have bright yellow-orange neck, breast, and bill patches;
incubate by placing their eggs on their feet, and when they hatch the chicks
are almost naked. This genus has a distribution centered on the Antarctic
coasts and barely extends to some subantarctic islands today.

Pygoscelis adeliae


Pygoscelis
adeliae
Picture:
© Samuel Blanc / www.sblanc.com
Pygoscelis contains species with a fairly simple black-and-white head pattern; their distribution is intermediate, centered on Antarctic coasts but extending somewhat northwards from there. In external morphology, these apparently still resemble the common ancestor of the Spheniscinae, as Aptenodytes' autapomorphies are in most cases fairly pronounced adaptations related to that genus' extreme habitat conditions. As the former genus, Pygoscelis seems to have diverged during the Bartonian, but the range expansion and radiation which led to the present-day diversity probably did not occur until much later; around the Burdigalian stage of the Early Miocene, roughly 20–15 mya.
The genera Spheniscus and Eudyptula contain species with a mostly subantarctic distribution centered on South America; some, however, range quite far northwards. They all lack carotenoid coloration, and the former genus has a conspicuous banded head pattern; they are unique among living penguins by nesting in burrows. This group probably radiated eastwards with the Antarctic Circumpolar Current out of the ancestral range of modern penguins throughout the Chattian (Late Oligocene), starting approximately 28 mya. While the two genera separated during this time, the present-day diversity is the result of a Pliocene radiation, taking place some 4–2 mya.

Pinguino
saltatore - Eudyptes
chrysocome
Picture:
© Samuel Blanc / www.sblanc.com
Pinguino saltatore - Eudyptes chrysocome
The Megadyptes–Eudyptes clade occurs at similar latitudes (though not as far north as the Galapagos Penguin), has its highest diversity in the New Zealand region, and represent a westward dispersal. They are characterized by hairy yellow ornamental head feathers; their bills are at least partly red. These two genera diverged apparently in the Middle Miocene (Langhian, roughly 15–14 mya), but again, the living species of Eudyptes are the product of a later radiation, stretching from about the late Tortonian (Late Miocene, 8 mya) to the end of the Pliocene.
The geographical and temporal pattern or spheniscine evolution corresponds closely to two episodes of global cooling documented in the paleoclimatic record. The emergence of the subantarctic lineage at the end of the Bartonian corresponds with the onset of the slow period of cooling that eventually led to the ice ages some 35 million years later. With habitat on the Antarctic coasts declining, by the Priabonian more hospitable conditions for most penguins existed in the subantarctic regions rather than in Antarctica itself. Notably, the cold Antarctic Circumpolar Current also started as a continuous circumpolar flow only around 30 mya, on the one hand forcing the Antarctic cooling, and on the other facilitating the eastward expansion of Spheniscus to South America and eventually beyond.
Later, an interspersed period of slight warming was ended by the Middle Miocene Climate Transition, a sharp drop in global average temperature from 14–12 mya, and similar abrupt cooling events followed at 8 mya and 4 mya; by the end of the Tortonian, the Antarctic ice sheet was already much like today in volume and extent. The emergence of most of today's subantarctic penguin species almost certainly was caused by this sequence of Neogene climate shifts.
Penguin ancestry beyond Waimanu remains unknown and not well-resolved by molecular or morphological analyses. The latter tend to be confounded by the strong adaptive autapomorphies of the Sphenisciformes; a sometimes perceived fairly close relationship between penguins and grebes is almost certainly an error based on both groups' strong diving adaptations, which are homoplasies. On the other hand, different DNA sequence datasets do not agree in detail with each other either.
The penguin is an accomplished swimmer, having flippers instead of wings. What seems clear is that penguins belong to a clade of Neoaves (living birds except paleognaths and fowl) which comprises what is sometimes called "higher waterbirds" to distinguish them from the more ancient waterfowl. This group contains such birds as storks, rails, and the seabirds, with the possible exception of the Charadriiformes.
Inside this group, penguin relationships are far less clear. Depending on the analysis and dataset, a close relationship to Ciconiiformes or to Procellariiformes has been suggested. Some think the penguin-like plotopterids (usually considered relatives of anhingas and cormorants) may actually be a sister group of the penguins, and that penguins may have ultimately shared a common ancestor with the Pelecaniformes and consequently would have to be included in that order, or that the plotopterids were not as close to other pelecaniforms as generally assumed, which would necessitate splitting the traditional Pelecaniformes in three.
Penguins are superbly adapted to an aquatic life. Their vestigial wings have become flippers, useless for flight in the air. In the water, however, penguins are astonishingly agile. Within the smooth plumage a layer of air is preserved, ensuring buoyancy. The air layer also helps insulate the birds in cold waters. On land, penguins use their tails and wings to maintain balance for their upright stance.
All penguins are countershaded for camouflage – that is, they have a white underside and a dark (mostly black) upperside. A predator looking up from below (such as an orca or a leopard seal) has difficulty distinguishing between a white penguin belly and the reflective water surface. The dark plumage on their backs camouflages them from above.
Diving penguins reach 6 to 12 km/h (3.7 to 7.5 mph), though there are reports
of velocities of 27 km/h (17 mph) (which are more realistic in the case of
startled flight). The small penguins do not usually dive deep; they catch
their prey near the surface in dives that normally last only one or two
minutes. Larger penguins can dive deep in case of need. Dives of the large
Emperor Penguin have been recorded which reach a depth of 565 m (1,870 ft) and
last up to 22 minutes.
Penguins either waddle on their feet or slide on their bellies across the snow,
a movement called "tobogganing", which conserves energy while moving
quickly. They also jump with both feet together if they want to move more
quickly or cross steep or rocky terrain.
Penguins have an average sense of hearing for birds; this is used by parents and chicks to locate one another in crowded colonies. Their eyes are adapted for underwater vision, and are their primary means of locating prey and avoiding predators; in air it has been suggested that they are nearsighted, although research has not supported this hypothesis.
Penguins have a thick layer of insulating feathers which serve to keep them warm in water (heat loss in water is much greater than in air). The Emperor penguin (the largest penguin) has the largest body mass of all penguins, which further reduces relative surface area and heat loss. They also are able to control blood flow to their extremities, reducing the amount of blood which gets cold, but still keeping the extremities from freezing. In the extreme cold of the Antarctic winter, the females are at sea fishing for food leaving the males to brave the weather by themselves. They often huddle together to keep warm and rotate positions to make sure that each penguin gets a turn in the center of the heat pack. They can drink salt water because their supraorbital gland filters excess salt from the bloodstream. The salt is excreted in a concentrated fluid from the nasal passages.
The Auk of the Northern Hemisphere is superficially similar to penguins: they are not related to the penguins at all, but considered by some[who?] to be a product of moderate convergent evolution.
Isabelline penguins - Perhaps one in 50,000 penguins (of most species) are born with brown rather than black plumage. These are called isabelline penguins, possibly in reference to the legend that the archduchess Isabella of Austria vowed not to change her undergarments until her husband united the northern and southern Low Countries by taking the city of Ostend – which took three years to accomplish. Isabellinism is different from albinism. Isabelline penguins tend to live shorter lives than normal penguins, as they are not well-camouflaged against the deep, and are often passed over as mates.
Although all penguin species are native to the southern hemisphere, they are not found only in cold climates, such as Antarctica. In fact, only a few species of penguin actually live so far south. At least 10 species live in the temperate zone; one, the Galápagos Penguin, lives as far north as the Galápagos Islands, but this is only made possible by the cold, rich waters of the Antarctic Humboldt Current which flows around these islands. Major populations of penguins are found in: Antarctica, Australia, New Zealand.
Penguins for the most part breed in large colonies, the exceptions being the Yellow-eyed and Fiordland species; these colonies may range in size from as few as a 100 pairs for Gentoo Penguins, to several hundred thousand in the case of King, Macaroni and Chinstrap Penguins. Living in colonies results in a high level of social interaction between birds, which has led to a large repertoire of visual as well as vocal displays in all penguin species. Agonistic displays are those which are intended to confront or drive off, or alternately appease and avoid conflict with, other individuals.
Penguins form monogamous pairs for a breeding season, though the rate the same pair recouples varies drastically. Most penguins lay two eggs in a clutch, although the two largest species, the Emperor and the King Penguins, lay only one. With the exception of the Emperor Penguin, in all penguins share the incubation duties. These incubation shifts can last days and even weeks as one member of the pair feeds at sea. Penguins generally only lay one brood; the exception is the Little Penguin which can raise two or three broods in a season.
Penguin eggs are smaller than any other bird species when compared proportionally to the weight of the parent birds; at 52 g (2 oz), the Little Penguin egg is 4.7% of its mothers' weight, and the 450 g (1 lb) Emperor Penguin egg is 2.3%. The relatively thick shell forms between 10 and 16 % of the weight of a penguin egg, presumably to minimise risk of breakage in an adverse nesting environment. The yolk, too, is large, and comprises 22–31 % of the egg. Some yolk often remains when a chick is born, and is thought to help sustain it if parents are delayed in returning with food.
When mothers lose a chick, they sometimes attempt to "steal" another mother's chick, usually unsuccessfully as other females in the vicinity assist the defending mother in keeping her chick. In some species, such as Emperor Penguins, young penguins assemble in large groups called crčches.
Penguins seem to have no special fear of humans, and have approached groups of explorers without hesitation. This is probably because penguins have no land predators in Antarctica or the nearby offshore islands. Instead, penguins are at risk at sea from predators such as the leopard seal. Typically, penguins do not approach closer than about 3 meters (10 ft) at which point they become nervous. This is also the distance that Antarctic tourists are told to keep from penguins (tourists are not supposed to approach closer than 3 meters, but are not expected to withdraw if the penguins come closer).
Penguins are popular around the world, primarily for their unusually upright, waddling gait and (compared to other birds) lack of fear of humans. Their striking black-and-white plumage is often likened to a tuxedo suit. Mistakenly, some artists and writers have penguins based at the North Pole. This is incorrect, as there are almost no wild penguins in the northern hemisphere, except the small group on the northernmost of the Galápagos. The cartoon series Chilly Willy helped perpetuate this myth, as the title penguin would interact with northern-hemisphere species such as polar bears and walruses.
Emperor Penguin
Aptenodytes forsteri
Class: Aves
Order: Sphenisciformes
Family: Spheniscidae
Genus: Aptenodytes
Species: forsteri, Gray, 1844
Picture:
© Samuel Blanc / www.sblanc.com
The Emperor Penguin (Aptenodytes forsteri) is the tallest and heaviest of all living penguin species and is endemic to Antarctica. The male and female are similar in plumage and size, reaching 122 cm (48 in) in height and weighing anywhere from 22–37 kg (48–82 lb). The dorsal parts are black and sharply delineated from the white belly, pale-yellow breast and bright-yellow ear patches. Like all penguins, it is flightless, with a streamlined body and wings stiffened and flattened into flippers for a marine lifestyle.
Fish form the bulk of its diet, which can include crustaceans, such as krill, and cephalopods, such as squid. In hunting, the species can remain submerged up to 18 minutes, diving to a depth of 535 m (1,755 ft). It has several adaptations to facilitate this, including an unusually structured haemoglobin to allow it to function at low oxygen levels, solid bones to reduce barotrauma, and the ability to reduce its metabolism and shut down non-essential organ functions.
The Emperor Penguin is perhaps best known for the sequence of journeys adults make each year in order to mate and to feed their offspring. The only penguin species that breeds during the Antarctic winter, it treks 50–120 km (31–75 mi) over the ice to breeding colonies which may include thousands of individuals. The female lays a single egg, which is incubated by the male while the female returns to the sea to forage; parents subsequently take turns foraging at sea and caring for their chick in the colony. The lifespan is typically 20 years in the wild, although observations suggest that some individuals may live to 50 years of age.
The Emperor Penguin was described in 1844 by English zoologist George Robert Gray, who made up its generic name from Ancient Greek word elements, a-ptēno-dytēs, "without-wings-diver". Its specific epithet is in honour of the German naturalist Johann Reinhold Forster, who accompanied Captain James Cook on his second Pacific Voyage and officially named five other penguin species.
Together with the similarly coloured but smaller King Penguin (A. patagonicus), the Emperor Penguin is one of two extant species in the genus Aptenodytes. Fossil evidence of a third species — Ridgen's Penguin (A. ridgeni) — has been found in fossil records from the late Pliocene, about three million years ago, in New Zealand. Studies of penguin behaviour and genetics have proposed that the genus Aptenodytes is basal; in other words, that it split off from a branch which led to all other living penguin species. Mitochondrial and nuclear DNA evidence suggests this split occurred around 40 million years ago.
The adult Emperor Penguin stands up to 122 cm (48 in) tall and can weigh from 22 to 45 kg (50–100 lb), depending on where it is in the reproductive cycle; both male and female penguins lose substantial mass while raising hatchlings and incubating eggs. Like all penguin species, it has a streamlined body to minimise drag while swimming, and wings that have become stiff, flat flippers. The tongue is equipped with rear-facing barbs to prevent prey from escaping when caught. Males and females are similar in size and colouration. The adult has deep black dorsal feathers, covering the head, chin, throat, back, dorsal part of the flippers, and tail. The black plumage is sharply delineated from the light-coloured plumage elsewhere. The underparts of the wings and belly are white, becoming pale yellow in the upper breast, while the ear patches are bright yellow. The upper mandible of the 8 cm (3 in) long bill is black, and the lower mandible can be pink, orange or lilac. In juveniles, the auricular patches, chin and throat are white, while its bill is black. The Emperor Penguin chick is typically covered with silver-grey down and has a black head and white mask. A chick with all-white plumage was found in 2001, but was not considered to be an albino as it did not have pink eyes. Chicks weigh around 315 g (11 oz) after hatching, and fledge when they reach about 50% of adult weight.
The Emperor Penguin's dark plumage fades to brown from November to February, before the yearly moult in January and February. Moulting is rapid in this species compared with other birds, taking only around 34 days. Emperor Penguin feathers emerge from the skin after they have grown to a third of their total length, and before old feathers are lost, to help reduce heat loss. New feathers then push out the old ones before finishing their growth.
The average yearly survival rate of the Emperor Penguin has been measured at
95.1%, with an average life expectancy of 19.9 years. The same researchers
estimated that 1% of Emperor Penguins born could feasibly reach an age of 50
years. In contrast, only 19% of chicks survive their first year of life.
Therefore, 80% of the Emperor Penguin population comprises adults five years
and older.
As the species has no fixed nest sites that individuals can use to locate
their own partner or chick, the Emperor Penguin must rely on vocal calls alone
for identification. It uses a complex set of calls that are critical to
individual recognition between parents, offspring, and mates, displaying the
widest variation in individual calls of all penguins. Vocalizing Emperor
Penguins use two frequency bands simultaneously. Chicks use a
frequency-modulated whistle to beg for food and to contact parents.
An Emperor Penguin can hold its breath anywhere up to 20 minutes, and dive over 550 meters (1,800 ft) The Emperor Penguin breeds in the coldest environment of any bird species; air temperatures may reach −40 °C (−40 °F), and wind speeds may reach 144 km/h (89 mph). Water temperature is a frigid −1.8 °C (28.8 °F), which is much lower than the Emperor Penguin's average body temperature of 39 °C (102 °F). The species has adapted in several ways to counteract heat loss. Feathers provide 80–90% of its insulation, and it has a layer of sub-dermal fat which may be up to 3 cm (1.2 in) thick before breeding. This resultant blubber layer impedes the mobility of the Emperor on land compared to its less well fat-insulated cousin, the Magellanic Penguin. Its stiff feathers are short, lanceolate (spear-shaped), and densely packed over the entire skin surface. With around 100 feathers covering one square inch (15/cm2), it has the highest feather density of any bird species. An extra layer of insulation is formed by separate shafts of downy filaments between feathers and skin. Muscles allow the feathers to be held erect on land, reducing heat loss by trapping a layer of air next to the skin. Conversely, the plumage is flattened in water, thus waterproofing the skin and the downy underlayer. Preening is vital in facilitating insulation and in keeping the plumage oily and water-repellent.
The Emperor Penguin is able to thermoregulate (maintain its core body temperature) without altering its metabolism, over a wide range of temperatures. Known as the thermoneutral range, this extends from −10 to 20 °C (10 to 70 °F). Below this temperature range, its metabolic rate increases significantly, although an individual can maintain its core temperature between 37.6 and 38.0 °C (99.7 to 100.4 °F) down to −47 °C (−53 °F). Movement by swimming, walking, and shivering are three mechanisms for increasing metabolism; a fourth process involves an increase in the breakdown of fats by enzymes, which is induced by the hormone glucagon. At temperatures above 20 °C (68 °F), an Emperor Penguin may become agitated as its body temperature and metabolic rate rise to increase heat loss. Raising its wings and exposing the undersides increases the exposure of its body surface to the air by 16%, facilitating further heat loss.
In addition to the cold, the Emperor Penguin encounters another adverse environment on deep dives — that of the markedly increased pressure of up to 40 times that of the surface, which in most other terrestrial organisms would cause barotrauma. The bones of the penguin are solid rather than air-filled, which eliminates the risk of mechanical barotrauma.
However, it is unknown how the species avoids the effects of nitrogen-induced decompression sickness. Oxygen use is markedly reduced, as its heart rate is reduced to as low as five beats per minute and non-essential organs are shut down, thus facilitating longer dives. The Emperor Penguin's haemoglobin and myoglobin are able to bind and transport oxygen at low blood concentrations; this allows the bird to function with very low oxygen levels that would normally result in the loss of consciousness.
The Emperor Penguin has a circumpolar distribution in the Antarctic almost exclusively between the 66° and 77° south latitudes. It almost always breeds on stable pack ice near the coast and up to 18 km (11 mi) offshore. Breeding colonies are usually located in areas where ice cliffs and icebergs shelter them from the wind. The total population is estimated at around 400,000–450,000 individuals, which are distributed among as many as 40 independent colonies. Around 80,000 pairs breed in the Ross Sea sector. Major breeding colonies are located at Cape Washington (20,000–25,000 pairs), Coulman Island in Victoria Land (around 22,000 pairs), Halley Bay, Coats Land (14,300–31,400 pairs), and Atka Bay in Queen Maud Land (16,000 pairs). Two land colonies have been reported: one on a shingle spit at Dion Island on the Antarctic Peninsula, and one on a headland at Taylor Glacier in the Australian Antarctic Territory. Vagrants have been recorded on Heard Island, South Georgia, and in New Zealand.
The Emperor Penguin is listed as a species of "least concern" by the IUCN. Along with nine other species of penguin, it is currently under consideration for inclusion under the US Endangered Species Act. The primary reasons for this are declining food availability due to the effects of climate change and industrial fisheries on the crustacean and fish populations. Other reasons for their potential placement on this list include disease, habitat destruction, and disturbance at breeding colonies by humans. Of particular concern is the impact of tourism. One study has shown Emperor Penguin chicks in a créche to become more apprehensive following helicopter approach to 1,000 m (3,281 ft). Population declines of 50% in the Terre Adélie region have been observed due to increased adult mortality, especially of males, during an abnormally prolonged warm period in the late 1970s, which resulted in reduced sea-ice coverage. On the other hand, egg hatching success rates declined when the sea-ice extent increased. The species is therefore considered to be highly sensitive to climatic changes.
A Woods Hole Oceanographic Institution study in January 2009 found Emperor Penguins could be pushed to the brink of extinction by the year 2100 due to global climate change. By applying mathematical models to predict how the loss of sea ice from climate warming would affect a big colony of Emperor Penguins at Terre Adélie, Antartica, they forecast a decline of 87% in the colony's population by the end of the century, from the current 3,000 breeding pairs in the colony to 400 breeding pairs. The decline may be mirrored in the whole Emperor Penguin population, estimated at about 200,000 breeding pairs. In 2009, satellite images of areas of excrement stained ice that are large enough to be visible from space helped scientists to discover ten previously unknown emperor penguin colonies in Antarctica.
The Emperor Penguin is a social animal in its nesting and its foraging behaviour; birds hunting together may coordinate their diving and surfacing. Individuals may be active day or night. A mature adult travels throughout most of the year between the nesting area and ocean foraging areas; the species disperses into the oceans from January to March.
The American physiologist Gerry Kooyman revolutionized the study of penguin foraging behaviour in 1971 when he published his results from attaching automatic dive-recording devices to Emperor Penguins. He found that the species reaches depths of 265 m (869 ft), with dive periods of up to 18 minutes. Later research revealed a small female had dived to a depth of 535 m (1,755 ft) near McMurdo Sound. It is possible that the Emperor Penguin can dive even deeper, as the accuracy of the recording devices is diminished at greater depths. Further study of one bird's diving behaviour revealed regular dives to 150 m (490 ft) in water around 900 m (3,000 ft) deep, and shallow dives of less than 50 m (160 ft), interspersed with deep dives of more than 400 m (1,300 ft) in depths of 450 to 500 m (1500 to 1600 ft). This was suggestive of feeding near or at the sea bottom.
Both male and female Emperor Penguins forage for food up to 500 km (311 mi) from colonies while collecting food to feed chicks, covering 82–1,454 km (51–904 mi) per individual per trip. A male returning to the sea after incubation heads directly out to areas of permanent open water, known as polynyas, around 100 km (62 mi) from the colony.
An efficient swimmer, the Emperor Penguin exerts pressure with both its upward and downward strokes while swimming. The upward stroke works against buoyancy and helps maintain depth. Its average swimming speed is 6–9 km/h (4–6 mph). On land, the Emperor Penguin alternates between walking with a wobbling gait and tobogganing — sliding over the ice on its belly, propelled by its feet and wing-like flippers. Like all penguins, it is flightless.
As a defence against the cold, a colony of Emperor Penguins forms a compact
huddle (also known as the turtle formation) ranging in size from ten to
several hundred birds, with each bird leaning forward on a neighbour. Those on
the outside tend to shuffle slowly around the edge of the formation, producing
a slow churning action, and giving each bird a turn on the inside and the
outside.
The Emperor Penguin's diet consists mainly of fish, crustaceans and
cephalopods, although its composition varies from population to population.
Fish are usually the most important food source, and the Antarctic silverfish
(Pleuragramma antarcticum) makes up the bulk of the bird's diet. Other
prey commonly recorded include other fish of the family Nototheniidae,
the Glacial Squid (Psychroteuthis glacialis), and the hooked squid
species Kondakovia longimana, as well as Antarctic krill (Euphausia
superba). The Emperor Penguin searches for prey in the open water of the
Southern Ocean, in either ice-free areas of open water or tidal cracks in pack
ice. One of its feeding strategies is to dive to around 50 m (164 ft), where
it can easily spot sub-ice fish like the Bald notothen (Pagothenia
borchgrevinki) swimming against the bottom surface of the sea-ice; it
swims up to the bottom of the ice and catches the fish. It then dives again
and repeats the sequence about half a dozen times before surfacing to breathe.
The Emperor Penguin's predators include birds and aquatic mammals; the Southern Giant Petrel (Macronectes giganteus) is the predominant avian predator, responsible for up to 34% of chick deaths in some colonies. The South Polar Skua (Stercorarius maccormicki) mainly scavenges for dead chicks, as the live chicks are too large to be attacked by the time of its annual arrival in the colony. The primary aquatic predators are both mammals: the Leopard Seal (Hydrurga leptonyx), which takes some adult birds, as well as fledglings soon after they enter the water, and the Orca (Orcinus orca), which takes adult birds.
The Emperor Penguin is able to breed at around three years of age, and usually commences breeding around one to three years later. The yearly reproductive cycle begins at the start of the Antarctic winter, in March and April, when all mature Emperor Penguins travel to colonial nesting areas, often walking 50 to 120 km (30 to 75 mi) inland from the edge of the pack ice. The start of travel appears to be triggered by decreasing day lengths; Emperor Penguins in captivity have been induced successfully into breeding by using lighting systems mimicking seasonal Antarctic day lengths. The penguins start courtship in March or April, when the temperature can be as low as −40 °C (−40 °F). A lone male gives an ecstatic display, where it stands still and places its head on its chest before inhaling and giving a courtship call for 1–2 seconds; it then moves around the colony and repeats the call. A male and female then stand face to face, with one extending its head and neck up and the other mirroring it; they both hold this posture for several minutes. Once in pairs, couples waddle around the colony together, with the female usually following the male. Before copulation, one bird bows deeply to its mate, its bill pointed close to the ground, and its mate then does the same.
Emperor Penguins are serially monogamous. They have only one mate each year, and stay faithful to that mate. However, fidelity between years is only about 15%. The narrow window of opportunity available for mating appears to be an influence, as there is a priority to mate and breed which often precludes waiting for the appearance of the previous year's partner.
The egg of the Emperor Penguin
It is 12 × 8 cm and vaguely pear-shaped
The female penguin lays one 460–470 g (1 lb) egg in May or early June; it is vaguely pear-shaped, pale greenish-white, and measures around 12 × 8 cm (4ľ x 3 in). It represents just 2.3% of its mother's body weight, making it one of the smallest eggs relative to the maternal weight in any bird species. 15.7% of the weight of an Emperor Penguin egg is shell; like those of other penguin species, the shell is relatively thick to minimise risk of breakage.
After laying, the mother's nutritional reserves are exhausted and she very carefully transfers the egg to the male, before immediately returning to the sea for two months to feed. The transfer of the egg can be awkward and difficult, and many couples drop the egg in the process. When this happens, the chick inside is quickly lost, as the egg cannot withstand the freezing temperatures on the icy ground. The male spends the winter incubating the egg in his brood pouch, balancing it on the tops of his feet, for 64 consecutive days until hatching. The Emperor Penguin is the only species where this behaviour is observed; in all other penguin species both parents take shifts incubating. By the time the egg hatches, the male will have fasted for around 115 days since arriving at the colony. To survive the cold and winds of up to 200 km/h (120 mph), the males huddle together, taking turns in the middle of the huddle. They have also been observed with their backs to the wind to conserve body heat. In the four months of travel, courtship, and incubation, the male may lose as much as 20 kg (44 lb), from around 38 kg to just 18 kg (84 lb to 40 lb).
Hatching may take as long as two or three days to complete, as the shell of
the egg is thick. Newly hatched chicks are semi-altricial, covered with only a
thin layer of down and entirely dependent on their parents for food and warmth.
If the chick hatches before the mother's return, the father feeds it a
curd-like substance composed of 59% protein and 28% lipid, which is produced
by a gland in his esophagus. The young chick is brooded in what is called the
guard phase, spending time balanced on its parent's feet and sheltered in the
brood pouch.
The female penguin returns at any time from hatching to ten days afterwards,
from mid-July to early August. She finds her mate among the hundreds of
fathers by his vocal call and takes over caring for the chick, feeding it by
regurgitating the food that she has stored in her stomach. The male then
leaves to take his turn at sea, spending around 24 days there before returning.
His trip is slightly shorter than it was originally, because the melting of
ice in the summer gradually decreases the distance between the breeding site
and the open sea. The parents then take turns, one brooding while the other
forages at sea.
About 45–50 days after hatching, the chicks form a crčche, huddling together for warmth and protection. During this time, both parents forage at sea and return periodically to feed their chicks. A crčche may comprise up to several thousand birds densely packed together and is essential for surviving the low Antarctic temperatures. From early November, chicks begin moulting into juvenile plumage, which takes up to two months and is often not completed by the time they leave the colony; adults cease feeding them during this time. All birds make the considerably shorter trek to the sea in December or January and spend the rest of the summer feeding there.
![]()