Vol. 2° - XII.2.
IL DIFFERENZIAMENTO cellulare e tissutale
Il fenomeno per cui da una
cellula iniziale, di solito lo zigote, si originano gradualmente tipi
cellulari diversi, seppure muniti dello stesso genoma, č detto
differenziamento. Alla base del differenziamento si trova una
particolare programmazione dell’attivitŕ genetica
che
in un primo tempo determina la formazione di tipi cellulari differenti, quindi
li mette in grado di rimanere specializzati, mantenendo costantemente represse
le sintesi che non sono specifiche di un determinato tipo cellulare.
La possibilitŕ di modulare nel
tempo e nello spazio l’attivitŕ dei vari gruppi di geni nucleari č stata
probabilmente alla base della grande fioritura evolutiva degli eucarioti,
molti dei quali, grazie a tale possibilitŕ, sono divenuti pluricellulari.
Un batterio, il cui genoma č continuamente attivo, dŕ
origine solo a cellule sempre eguali fra loro, che non traggono vantaggi dall’associazione
perché tutte esercitano le stesse attivitŕ.
L’organismo dei pluricellulari č fatto da tipi
cellulari diversi i quali, modulando in modo diverso l’attivitŕ di singoli
gruppi genici, si specializzano ciascuno per determinate funzioni e quindi
ricavano vantaggi dalla collaborazione reciproca all’interno di un unico
individuo.
In un organismo adulto le cellule dei vari tessuti
differiscono tra loro per forma e funzioni. A livello molecolare tali
differenze riguardano soprattutto proteine di carattere strutturale o
enzimatico. La fibra muscolare č enormemente piů ricca di mioglobina,
miosina, actina ed altre proteine contrattili di qualsiasi altra cellula,
mentre un globulo rosso non produce altro che emoglobina.
Alla base del differenziamento c’č quindi una particolare programmazione dell’attivitŕ
genetica, che prima determina la formazione di tipi cellulari
differenti, quindi li mette in grado di rimanere specializzati, mantenendo
costantemente represse le sintesi che non sono specifiche di quel tipo
cellulare.
Il
nucleo non č il solo responsabile di questa programmazione:
infatti, le cellule dei vari tessuti di un organismo hanno tutte lo stesso
DNA. La diversa regolazione dell’attivitŕ genetica che si attua nei vari
tipi cellulari dipende da segnali chimici che giungono al nucleo dal
citoplasma, oppure, sempre mediati dal citoplasma, da cellule circostanti o
anche dall’ambiente esterno alla cellula.
Segnali
chimici analoghi sono, del resto, prodotti anche nel corso della
vita cellulare, indipendentemente dal differenziamento: i complessi cicli
vitali della cellula eucariotica sono programmati
correttamente da molecole specifiche che vengono sintetizzate in determinati
momenti e inducono il nucleo a iniziare una nuova fase di attivitŕ. Le interazioni nucleo-citoplasmatiche
sono quindi alla base sia del differenziamento sia della normale attivitŕ di
una cellula nel corso del suo ciclo vitale. Vediamo alcuni esempi di questi
fenomeni, che di norma si svolgono nella fase G1 ma che possono interessare
anche periodi successivi.
La nascita dei vari tipi cellulari nel corso dello sviluppo dipende dalla struttura molecolare della cellula-uovo. Fin dalla fecondazione, o anche prima, in alcuni organismi si puň vedere che il citoplasma ovulare non č omogeneo per quanto riguarda la distribuzione di determinati costituenti, e in particolare delle ribonucleoproteine. Si tratta, probabilmente, di RNA messaggeri temporaneamente inattivi in quanto bloccati dagli informosomi.
Man
mano che l’uovo si va segmentando in numerosi blastomeri, che sono cellule
ancora indifferenziate, ciascun
blastomero eredita una diversa porzione del citoplasma ovulare
e quindi delle RNP differenti. Piů tardi, dai blastomeri si originano cellule
che si avviano in varie direzioni differenziatrici: alcune divengono
muscolari, altre nervose, altre intestinali e cosě via. Si ritiene che tale
processo sia dovuto a una diversa regolazione del loro DNA da parte di
proteine citoplasmatiche particolari, costruite sui vari mRNA derivati dall’uovo
e ora attivati.
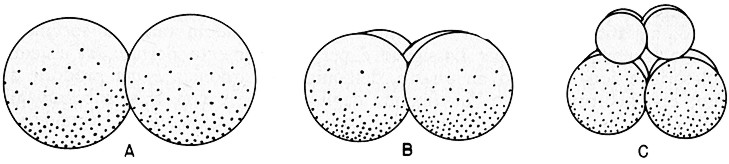
Fig. XII. 1 - Segmentazione e differenziamento. L’immagine schematica vuole sottolineare un fatto: con il moltiplicarsi dello zigote si realizza una differenza tra la metŕ superiore e inferiore del citoplasma. A stadio a due cellule. B stadio a quattro cellule. C stadio a otto cellule; le quattro cellule superiori sono giŕ differenziate rispetto alle quattro inferiori.
Tali controllori
citoplasmatici delle attivitŕ nucleari sono ancora sconosciuti. L’unico
noto č il fattore o+, ova-deficient,
scoperto nell’Axolotl, un anfibio che spesso non va incontro a metamorfosi.
Tale fattore č una proteina la cui assenza provoca la produzione di uova che
si arrestano nello sviluppo. Si ritiene che la proteina o+
sia necessaria per l’attivazione dei geni dei blastomeri, che altrimenti
restano repressi. Infatti, uova segmentate di individui deficienti di tale
fattore, destinate ad abortire, riprendono un normale sviluppo se viene loro
iniettata la proteina o+, ripresa che si manifesta con
un ripristino della trascrizione nei nuclei dei blastomeri.
La
segregazione in determinate aree dell’uovo di RNP particolari
č provata anche dalle esperienze sui plasmi
polari, zone ovulari ricche in granuli di RNA (granuli polari dalle quali
si originano normalmente le gonadi dell’individuo. Se nell’uovo di una
Drosofila vengono trapiantati granuli polari di un altro uovo, si origina un
embrione con un duplice
corredo di gonadi, uno derivato dal proprio plasma polare, l’altro
da quello trapiantato. Questi granuli contengono forse gli mRNA di proteine
che regolano il differenziamento delle cellule germinali.
Anche le esperienze di trapianto nucleare indicano che il
citoplasma contiene molecole in grado di modificare le attivitŕ nucleari.
Č noto che il genoma di cellule differenziate trascrive
pochi tipi di RNA, essendo in larga misura represso. Se il nucleo di una di
queste cellule č trapiantato in un uovo privato del proprio nucleo, nel nuovo ambiente il nucleo differenziato subisce una riprogrammazione:
il DNA inizia ampi fenomeni di trascrizione e il nucleo č in grado di
produrre il normale sviluppo dell’uovo, con la genesi di numerosi e diversi
tipi cellulari. Per esempio, in una specie di anfibi si sono ottenuti
individui completi iniettando nuclei di cellule intestinali in uova
previamente enucleate.
I fenomeni regolativi dell’attivitŕ genetica sono
dunque reversibili, in base a stimoli provenienti dal citoplasma. Un’ulteriore
prova si ha da esperienze di fusione cellulare o ibridazione
cellulare.
La fusione di due cellule genera un ibrido con due nuclei
in un unico citoplasma. Se le cellule appartengono a due specie diverse, l’ibrido
č detto eterocarione.
Sono stati ottenuti eterocarioni provocando la fusione di eritrociti di pollo,
il cui nucleo č del tutto eterocromatico e quindi represso, con fibroblasti
umani o di topo, il cui nucleo č invece molto attivo nella trascrizione. Nell’ibrido
si puň vedere che il nucleo dell’eritrocita rapidamente si ingrandisce e la
sua cromatina diviene attiva, trascrivendo RNA e fabbricando il nucleolo di
cui era privo. Evidentemente, fattori presenti nel citoplasma del fibroblasto
hanno attivato il DNA dell’eritrocita.
Infine, č stato dimostrato che gli ormoni steroidei
attivano alcuni tipi cellulari, quelli degli organi bersaglio, penetrando nel
loro citoplasma e legandosi a recettori specifici di natura proteica. Tali
recettori, attivati dallo steroide, passano nel nucleo ed entrano in rapporto
con determinate regioni del genoma sulle quali agiscono come derepressori,
attivandone la trascrizione. Prove al riguardo si sono ottenute per gli
steroidi sessuali (testosterone, estrogeni e progesterone) e della corteccia
del surrene nei vertebrati e per quelli della muta larvale negli Insetti
(ecdisone).
In quest’ultimo caso si č visto che nei cromosomi
politenici
[1]
dei ditteri l’iniezione di ecdisone provoca la formazione di puffs (aree despiralizzate ad intensa attivitŕ trascrizionale) in
determinate regioni di certi cromosomi che sono specifiche sia a livello di
tessuto sia a livello dello stadio larvale osservato. Probabilmente nei puffs
piů grandi, oltre a fenomeni di attivazione, ne esistono anche altri di
amplificazione dell’attivitŕ genica.
Amplificazione genica, ossia incremento numerico delle
copie di un certo gene in attivitŕ, č stata di recente indotta
sperimentalmente anche in cellule di mammiferi in coltura. Trattando queste
cellule con inibitori della sintesi delle purine, sintesi che č di vitale
importanza, alcune di esse acquisivano gradualmente una resistenza all’agente
inibitore, aumentando enormemente la quantitŕ degli enzimi addetti alla
sintesi delle purine, che quindi non venivano totalmente inibiti. A livello
microscopico si č visto che queste cellule resistenti possedevano un
cromosoma con un braccio enormemente piů lungo della norma: ci sono prove che
lungo tale braccio i geni che codificano l’enzima per la sintesi
purinica sono presenti in alcune centinaia di copie, essendosi amplificati
nel corso di varie generazioni, sotto la pressione selettiva degli inibitori.
I principali eventi che caratterizzano le attivitŕ vitali
della cellula sono dunque il risultato di interazioni talora complesse fra
nucleo, citoplasma e ambiente che circonda la cellula.
2.1. Differenziamento cellulare
Abbiamo visto come il problema
del differenziamento molecolare si identifichi con quello dell’espressione
del gene. Il passo successivo č quello di rispondere alla domanda di come
possa una cellula organizzare una struttura perfettamente in grado di
espletare funzioni articolate. Abbiamo buoni motivi per ritenere che questo si
realizzi grazie ad un ordinato montaggio di macromolecole proteiche, lipidiche
e zuccherine secondo un preciso progetto
strutturale che richiede l’intervento di attivitŕ enzimatiche in una
sequenza temporale estremamente precisa. Generalmente l’acquisizione di una determinata funzione da parte di una
popolazione cellulare si realizza quando in questa si č progressivamente
accumulata una quantitŕ considerevole di una o piů proteine specifiche.
Negli eucarioti le proteine sintetizzate possono essere
distinte in due classi:
o
proteine responsabili dell’impalcatura strutturale della cellula;
o
proteine enzimatiche caratteristiche, in grado di
espletare funzioni specializzate.
Una caratteristica importante di alcune proteine del
secondo gruppo č quella di non essere strettamente necessarie al metabolismo
e in particolare alla sopravvivenza della cellula stessa. Pertanto, ai fini
dell’espressione di attivitŕ cellulari caratteristiche, č necessario che
ogni singola cellula sintetizzi non soltanto proteine in quantitŕ
sufficiente, ma anche rispettando una rigida sequenza temporale.
Durante il differenziamento, la cellula, in base alla
qualitŕ e alla disponibilitŕ delle proteine messe in produzione, č in grado
successivamente di assemblarle acquisendo caratteristiche fenotipiche
fortemente differenziate. Queste modificazioni sono a carico di tutti i
compartimenti cellulari che possono tuttavia ulteriormente differenziarsi in
rapporto alle funzioni specifiche che la cellula dovrŕ svolgere nel corso del
suo stato di differenziamento. Č importante sottolineare come la funzione
finale svolta da ogni singola popolazione sia in gran parte dipendente dall’assortimento
e dal numero dei macchinari molecolari presenti all’interno della cellula.
In particolare lo
sviluppo dei vari compartimenti cellulari č responsabile di conferire talune
caratteristiche attivitŕ funzionali.
2.2. Variazioni ultrastrutturali dovute al differenziamento
Č interessante dare uno sguardo
alle differenze strutturali che i meccanismi differenziativi determinano a
carico di alcuni tra i piů importanti organuli cellulari.
2.2.a. La membrana plasmatica
Cellule che si sviluppano sulle
pareti delle cavitŕ di un organo, in cui operano attivamente, possono
specializzare la loro porzione apicale per poter lavorare in questa cavitŕ in
cui normalmente puň essere presente materiale macro e micromolecolare di
varia natura che deve essere assorbito, modificato o semplicemente analizzato.
La porzione apicale delle cellule epiteliali dell’intestino,
dette enterociti, riesce ad amplificare notevolmente la superficie cellulare
attraverso una ripetuta pieghettatura che conferisce un aspetto a pettine. Le
popolazioni cellulari che rivestono il lume della cistifellea presentano dei
microvilli con antenne,
presumibilmente espressioni differenziate del cell coat che favoriscono il riassorbimento dell’acqua, importante
ai fini della concentrazione della bile. Nella mucosa tracheale esistono
cellule dotate di ciglia che, contraendosi, svolgono un lavoro meccanico di
allontanamento nei confronti del materiale in transito.
Molte volte la presenza di ciglia puň addirittura
promuovere o indurre il movimento di popolazioni cellulari, come accade nei
condotti efferenti delle vie spermatiche maschili e degli elementi che
rivestono la mucosa delle tube uterine: grazie all’attivitŕ meccanica
esercitata dalle ciglia, che battono in maniera orientata, ritmica e precisa,
si creano dei moti propulsivi attivi in grado di determinare, rispettivamente,
lo spostamento delle cellule spermatiche e dell’ovocellula.
Un ultimo esempio di collegamento fra struttura e funzione
lo troviamo nei melanociti
i quali, attraverso le propaggini citoplasmatiche denominate dendriti, possono mettersi in contatto con le cellule dell’epidermide
per riversarvi il pigmento melanico.
Durante il differenziamento di queste popolazioni
cellulari risulta evidente come debba essere fortemente incrementata la
sintesi dei vari materiali molecolari che costituiscono queste strutture e
come debba essere particolarmente controllato il loro successivo montaggio.
Attraverso la comparsa di caratteristiche fenotipiche fortemente specializzate
le varie popolazioni cellulari riescono a svolgere funzioni diverse e
complesse.
2.2.b. Il reticolo endoplasmatico
L’estensione del reticolo
endoplasmatico rugoso č un criterio differenziativo in grado di fornire
importanti informazioni sulle capacitŕ di sintesi di una cellula. La cellula
pancreatica presente nell’acino esocrino č un esempio di come sia possibile
correlare una marcata attivitŕ secretoria con un imponente sviluppo del REG.
L’elaborato di questa cellula andrŕ a costituire il succo pancreatico,
miscela di una quarantina circa di enzimi, che verrŕ immessa nella cavitŕ
duodenale per integrare l’attivitŕ intestinale nella demolizione delle
molecole presenti nel chimo.
Il reticolo liscio dell’epatocita puň variamente
ampliarsi durante tutto il corso della vita in seguito all’azione di
sostanze eterogenee tra cui sicuramente le piů interessanti risultano i
farmaci. Č stato calcolato che, in seguito a somministrazione di barbiturici,
il reticolo liscio, per un fenomeno detto farmacoinduzione, puň
ipertrofizzarsi raggiungendo valori pari a circa 45.000 mm˛.
Il reticolo liscio č senza dubbio la struttura piů
enigmatica dello sviluppo in quanto non č possibile indirizzarsi sul tipo di
funzione svolta dalla cellula basandosi solamente sull’analisi della sua
organizzazione strutturale. Durante la citodifferenziazione il reticolo
liscio, scarso e poco delineato nelle cellule pluripotenti, tende a
svilupparsi costituendo complicati sistemi di tubuli e di cisterne, che
attraversano tutta la cellula interconnettendosi tra loro. Pur presentandosi
al microscopio elettronico con atteggiamenti strutturali simili, il reticolo
liscio puň espletare funzioni diverse
2.2.c. Il complesso di Golgi
Durante la citodifferenziazione
l’apparato di Golgi va incontro ad una serie di importanti modificazioni
strutturali che interessano sia la sua estensione che la dilatazione delle sue
cisterne.
Dall’analisi di vari tipi cellulari si deve dedurre che
l’evoluzione, il numero e l’ampiezza delle cisterne del complesso di Golgi
non sono sempre un indice dell’attivitŕ di tale apparato. Questo perché la
sua funzionalitŕ č strettamente dipendente dall’attivitŕ degli enzimi che
esso contiene. Pertanto la disponibilitŕ di substrati (proteine e zuccheri) e
dei sistemi enzimatici golgiani (glicosil-transferasi) č l’elemento chiave
dell’attivitŕ di tale apparato. Le dimensioni del complesso di Golgi non
sono pertanto sempre correlabili alla sua reale capacitŕ di arricchimento
della quota proteica sintetizzata e il dato strutturale, da solo, non č un
indice valido della sua citodifferenziazione.
2.2.d. I mitocondri
Poco si sa di come i mitocondri
vadano organizzandosi durante il differenziamento. In genere, in una cellula
in stato differenziato come l’epatocita, il loro numero oscilla intorno al
migliaio. Per una buona valutazione dell’attivitŕ mitocondriale durante il
differenziamento non sempre il numero č un criterio fondamentale. Infatti,
ugualmente importanti da un punto di vista morfologico, sono la grandezza del
mitocondrio e il numero delle creste.
Secondo valutazioni stereologiche, poco piů di un
migliaio di mitocondri presenta una superficie pari a circa 720 mm˛. Le creste mitocondriali
possono raggiungere uno sviluppo di circa 35.000 mm˛.
Č chiaro che non soltanto attraverso il numero dei mitocondri, quanto
soprattutto attraverso l’estensione delle creste, si puň pervenire ad una
precisa valutazione morfologica dell’attivitŕ mitocondriale. Ciononostante
č innegabile l’alto numero di espressioni morfologiche che si possono
osservare a carico del compartimento mitocondriale durante lo sviluppo e
durante il mantenimento dello stato differenziato.
Il tessuto adiposo bruno, scarso nell’uomo adulto ma
presente in gran quantitŕ durante lo sviluppo embrionale, caratteristico di
alcune specie che prevedono l’ibernazione nel loro ciclo vitale, presenta
dei mitocondri assai interessanti. A parte le notevoli dimensioni, mostrano un
marcato sviluppo delle creste e una stretta associazione con i depositi
lipidici coi quali interagiscono promovendo la decarbossilazione degli acidi
grassi.
2.3. Differenziamento tissutale
Col termine di istodifferenziamento
si intende la progressiva organizzazione in raggruppamenti delle varie
popolazioni cellulari, detti tessuti. Pertanto, un tessuto nasce dalla
confluenza di una o piů popolazioni cellulari che si organizzano per dar
luogo a complessi morfo-funzionali altamente specializzati.
Durante l’istodifferenziazione alcune popolazioni
cellulari possono riunirsi in comunitŕ organizzate a costituire gli epiteli.
Per poter raggiungere questi livelli di organizzazione sono necessarie due
importanti condizioni strutturali: la presenza di un supporto su cui
attaccarsi, la lamina basale, e la possibilitŕ di connettersi fisicamente e
di comunicare con le cellule adiacenti tramite complessi di giunzione.
Le cellule, durante il differenziamento, oltre che a
specializzare i vari compartimenti intracellulari per poter raggiungere l’organizzazione
tissutale, nell’ambito della loro porzione basale devono in via preliminare
organizzare specifiche classi di glicoproteine che assicurano l’adesione
alla lamina basale sottostante. Inoltre devono mettere in produzione sistemi
specifici di connessione, quali i desmosomi, che assicurano tenacemente l’adesione
meccanica alla cellula vicina. Accanto a sistemi di connessione č importante
che tutte le popolazioni cellulari che compongono lo stesso tessuto abbiano il
medesimo livello di informazione
metabolica. Ciň č di fatto assicurato dalla presenza di specifici canali
di comunicazione, quali le gap-junctions.
Si comprende facilmente come l’organizzazione di un determinato tipo di
comunitŕ epiteliale abbia il suo presupposto nel differenziamento dei
congegni che assicurano la possibilitŕ dell’organizzazione in comunitŕ.
Altre popolazioni cellulari, quasi esclusivamente quelle
di origine mesenchimale, non operano in raggruppamenti, bensě isolate nell’ambito
di una matrice di varia consistenza, da loro stesse prodotta e controllata. Si
vengono cosě a costituire i tessuti connettivi che adempiono al compito
fondamentale di connettere tra loro
gli altri tessuti. Anche in questo caso, l’acquisizione delle
caratteristiche tessutali specifiche dipende dalla sintesi di determinati
prodotti che le cellule devono necessariamente elaborare per poter raggiungere
un certo livello istodifferenziativo. Talune popolazioni cellulari possono
incrementare fortemente la sintesi dei materiali extracellulari, organizzando
cosě altri sistemi tessutali, come il tessuto cartilagineo e il tessuto
osseo.
Di origine mesenchimale sono le cellule ematiche sospese
nella matrice connettivale liquida, il plasma, a sua volta contenuto nel
sistema vascolare. Ciň, tuttavia, non impedisce ad alcune di queste cellule,
quali alcuni tipi di leucociti, di abbandonare la rete vascolare a livello
dell’endotelio dei capillari per raggiungere il connettivo, dove si
trasformano in cellule dotate di attivitŕ fagocitaria.
Infine, un ultimo livello di organizzazione tessutale č
dato dai neuroni, particolari cellule interconnesse tra loro attraverso
specifiche giunzioni, le sinapsi, costituendo cosě estese reti
pluricellulari. I neuroni si sono altamente differenziati per assolvere a due
peculiari funzioni: l’eccitabilitŕ e la conduttivitŕ.
La comparsa nel corso dello sviluppo embrionale di tessuti
con forme e strutture nuove č un processo che comporta una complessa
ridistribuzione cellulare grazie a proliferazioni
programmate. Le popolazioni cellulari in sviluppo si autocontrollano,
indicandosi a vicenda dove migrare, dove sistemarsi nel contesto di una
popolazione cellulare, che cosa sintetizzare, quando e con quali modalitŕ.
Quindi, in seguito al differenziamento istologico, si
formano specifici raggruppamenti di cellule altamente specializzate che
costituiscono i tessuti. Generalmente, per tessuto si intende un insieme di
cellule aventi la stessa forma, la stessa funzione e la stessa derivazione
embrionale. I tessuti vengono raggruppati in quattro grandi classi:
epiteliali, connettivali, muscolari e nervosi. All’interno di ciascuna
classe, perň, si annoverano numerose sottoclassi: si ritiene che, nell’organismo
umano, esistano circa 500 popolazioni cellulari.
I tessuti si associano tra loro, concorrendo alla
formazione degli organi, i quali, a loro volta, si associano a costituire
sistemi e apparati tra loro correlati per lo svolgimento delle funzioni vitali
di un organismo.
|
sommario |
avanti |
|
|
|