Lessico
La struttura dell'uovo
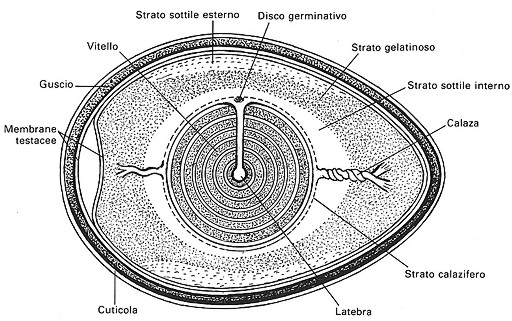
Disegno schematico
dell'uovo
che mette bene in evidenza la cuticola, il guscio e
le membrane testacee.
Nell'uovo deposto bisogna
distinguere:
À
la
cellula uovo, o tuorlo, o rosso d'uovo
À
lÆalbume,
o bianco d'uovo
À
il
guscio, che comprende anche le membrane testacee.
Anche se lÆuovo acquista la
forma e il volume caratteristici della specie, la proporzione relativa dei
costituenti Þ abbastanza costante.
|
Percentuale
dei componenti
da Botte & Pelagalli |
|||
|
Componenti |
Guscio |
Albume |
Cellula uovo |
|
acqua |
1 |
88,5 |
47,5 |
|
proteine |
4 |
10,5 |
17,4 |
|
lipidi |
|
|
33 |
|
carboidrati |
|
0,5 |
0,2 |
|
ioni
minerali |
95 |
0,5 |
1,1 |
|
altri |
|
|
0,8 |
|
|
|
|
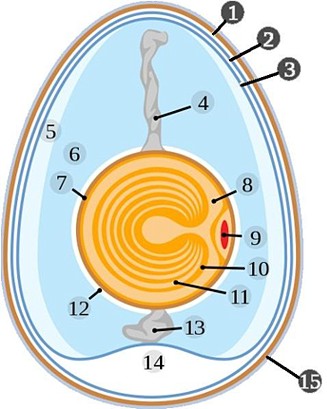
Cellula uovo
La cellula uovo ha forma
subsferica e in un uovo medio di 58 g
pesa circa 19 g con un diametro di 35-40
mm
e un volume di 16-17 cm│. Consta di tre parti: la membrana vitellina,
il disco germinativo e il vitello.
La
membrana vitellina avvolge il tuorlo, ma va distinta dalla struttura omonima
della cellula uovo contenuta nel follicolo ovarico. Di questa, infatti,
restano solo dei frammenti in quanto si dissolve in parte prima
dell'ovulazione. La membrana vitellina presenta una struttura complessa,
essendo formata da due strati di derivazione ovarica - i resti della membrana
vitellina e la membrana perivitellina - e da due strati probabilmente prodotti
dall'infundibolo e quindi derivati dall'albume: la membrana continua e la
membrana extravitellina.
Nell'ambito della membrana perivitellina
si osservano delle strutture fibrose ampiamente anastomizzate a rete e
orientate parallelamente alla superficie dellÆuovo spesse 2 mm.ÀLa membrana
continua, spessa 50-100 mm,
ha un aspetto granulare, mentre al suo esterno la membrana
extravitellina appare come una rete di sottili fibrille spessa 3-9
mm.
Nel suo complesso la membrana vitellina ha una notevole
resistenza, specialmente verso la regione appuntita dell'uovo. ╚ costituita
per l'80% da acqua; il 90% dei
solidi Þ rappresentato da proteine e il 3% da
lipidi. In linea di massima le sue parti pi¨ interne somigliano al collagene,
mentre quelle pi¨ esterne sono abbastanza simili all'albume.
La permeabilitÓ della membrana Þ alquanto
caratteristica, in quanto si lascia attraversare quasi esclusivamente da
acqua, che viene scambiata fra tuorlo e albume.
Il disco germinativo Þ una piccola parte
della cellula uovo che a causa della sua minore densitÓ occupa il polo
animale. Nell'uovo fecondato consta di una massa di 40.000-60.000 cellule
derivate dalla divisione ripetuta dello zigote e prende il nome di discoblastula
o blastoderma, che appare come un dischetto di colore grigio chiaro del
diametro di 4 mm che riposa sulla componente bianca del vitello. Nel
caso lÆuovo non sia stato fecondato, il disco germinativo Þ costituito da
citoplasma e dal nucleo femminile in degenerazione e il suo diametro Þ di
circa 3,5 mm.
Il vitello rappresenta la componente
maggiore della cellula uovo ed Þ strutturato in due porzioni facilmente
distinguibili. Al di sotto del disco germinativo si trova una struttura conica
di aspetto chiaro che si approfonda verso il centro della cellula, ove termina
con una porzione sferica di circa 5 mm
di diametro, la latÞbra,
composta di vitello bianco, e che rappresenta appena l'1-2%
del
totale. Attorno alla latebra si dispone il vitello giallo.
Si Þ molto discusso se questo sia presente in strati sovrapposti, dal momento
che in molti casi appare come un'alternanza di fasce giallo-rosse di diversa
intensitÓ. Oggi pare assodato che tale aspetto non rispecchi una particolare
disposizione spaziale, ma dipenda dalla ricchezza in xantofilla della dieta.
Albume
Anche lÆalbume presenta una
tipica organizzazione macroscopica in quanto non Þ fisicamente omogeneo. Si
distinguono due calaze e diversi strati.
Le calaze si dipartono da ciascun polo
della cellula uovo e sono dirette secondo lÆasse maggiore del guscio. Si
tratta di strutture cordoniformi avvolte su se stesse. Verso il polo ottuso si
dirige una sola calaza, mentre dall'altro lato ne esistono due tra loro
intimamente ritorte. Originano a livello dello strato calazifero e terminano
da ciascun lato nella regione dei legamenti dell'albume.
Lo strato calazifero Þ cospicuo, ha
struttura omogenea ed Þ a stretto contatto con la membrana vitellina della
cellula uovo.
Gli strati sottili interno ed esterno appaiono
piuttosto fluidi e omogenei; nella loro compagine non si distinguono fibre,
anche se sono presenti mucine.
Lo strato gelatinoso Þ molto ricco in
ovomucina che conferirebbe lÆaspetto di gel denso. A ciascuna estremitÓ
dell'asse maggiore dell'uovo si prolunga nei legamenti dell'albume connessi
mediante mucine alle membrane testacee. Si Þ molto discusso
sull'organizzazione submicroscopica di questa porzione dell'albume e
soprattutto Þ dibattuta la presenza o meno di fibre.
Membrane testacee
Si tratta di due membrane
leggermente elastiche e biancastre accollate fra loro, salvo che in
corrispondenza del polo ottuso dell'uovo dove formano la camera d'aria.
Quest'ultima si genera per il rapido raffreddamento dellÆuovo dopo la
deposizione, che porta alla penetrazione di aria e allo scollamento delle due
lamine. Ciascuna membrana Þ composta da pi¨ strati.
La membrana interna Þ organizzata in almeno
tre piani di fibre orientate in modo differente, per uno spessore totale di
circa 22 mm.
La membrana esterna Þ pi¨ complessa. Ha
uno spessore di circa 50 mm
e possiede fibre brevi e spesse, poste in almeno 6 piani
e orientate ad angolo retto tra piano e piano.
Guscio
Il guscio dellÆuovo Þ composto da pi¨ strati, che a partire dall'interno si susseguono nel seguente ordine:
À Membrane testacee: si tratta di due lamine tra loro accollate, applicate internamente al guscio calcareo, leggermente separate in corrispondenza del polo ottuso dell'uovo dove si trova la camera d'aria. Si tratta di strutture fibrose di tipo collagene, impropriamente omologate alla cheratina.
À Guscio propriamente detto: nel pollo ha uno spessore medio di circa 350 mm e contiene carbonato di calcio sotto forma di calcite, che ne rappresenta il 98%; il rimanente 2% Þ costituito da proteine; il guscio propriamente detto pu‗ essere ulteriormente suddiviso in due parti:
À
strato mammillare interno:
costituito da cristalli di calcite che hanno un diametro di 10-15 Ám
À strato spongioso esterno:
fatto anchÆesso di cristalli di calcite molto fini, che costituiscono i 2/3
del guscio.
À Cuticola: rappresenta lo strato pi¨ superficiale, dello spessore di circa 10 mm, formato da una particolare proteina analoga al collagene, la quale si continua sino allÆalbume attraversando i due primi strati, ma che Þ comunque diversa dalla piccola quota proteica del guscio.
L'osservazione al microscopio elettronico ha permesso di chiarire che la deposizione dei sali di calcio nella matrice del guscio prende lÆavvio da nuclei di cristallizzazione distribuiti sulla superficie esterna della membrana testacea superficiale, i cosiddetti core mammillari. Segue, quindi, un'apposizione di cristalli e di matrice in senso verticale con formazione di tante microcolonne che si associano tra loro, fatta eccezione per i punti in cui persistono delle discontinuitÓ - i pori - che nel pollo variano da 7.000 a 17.000. Una volta terminato questo processo, in superficie viene applicata la cuticola, in parte prodotta durante il rapido passaggio in vagina.
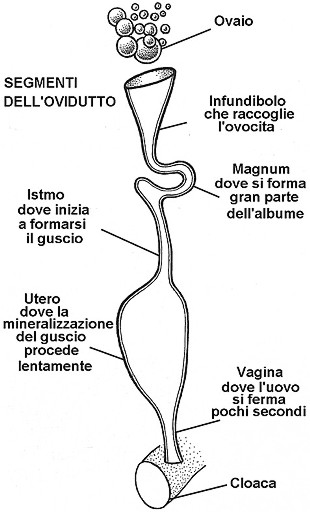
Ovidutto di gallina
Rappresentazione schematica
che non rispetta le effettive lunghezze dei
vari segmenti
Il guscio comincia a formarsi nell'istmo, ma inizia a delinearsi non appena lÆuovo giunge in utero. Pu‗ darsi che la sua produzione prosegua per tutto lÆarco di tempo che intercorre fra lÆinizio dellÆapposizione calcarea e il momento dellÆovodeposizione. Tuttavia i dati concordano sul fatto che lÆuovo Þ rigido giÓ 13 ore prima di essere deposto.
A livello dellÆistmo vengono ovviamente dapprima secrete le membrane testacee, dove fanno la loro comparsa i core mammillari, applicati alla pi¨ esterna di queste due membrane.
La mineralizzazione procede lentamente nell'utero, dove il pompaggio di acqua nell'uovo provvede a distenderlo, contribuendo forse alla formazione dei pori.
LÆintimo meccanismo responsabile della calcificazione Þ noto solo in parte; certamente i sali, sotto forma di ioni calcio e carbonato, sono sottratti dal sangue e vengono poi trasformati in calcite con il concorso sia delle ghiandole che delle cellule epiteliali della mucosa uterina.
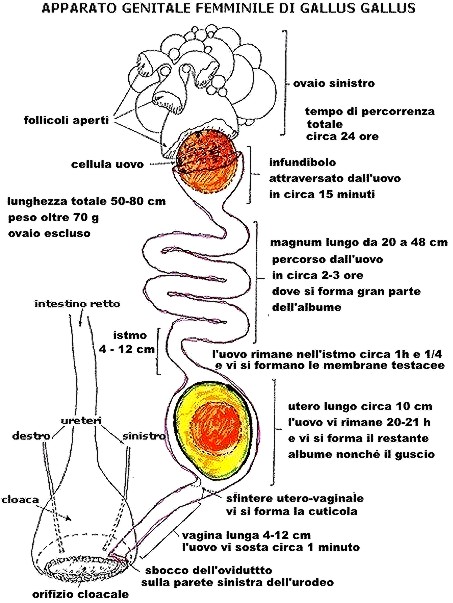
Schema
di ovidutto di gallina
Non
sono rispettate le reali lunghezze dei vari segmenti
Nucleus
of Pander![]()
Nucleo
di Pander
Il tuorlo bianco, al di sotto della membrane vitellina, avvolge tutta la superficie del tuorlo e, in corrispondenza del disco germinativo, forma una specie di cuscinetto noto come nucleo di Pander, da dove si spinge fino al centro del tuorlo costituendo la latebra di Purkinje. Al di sotto del primo strato bianco che circonda tutto il tuorlo ve ne Þ uno giallo a cui segue uno bianco e cosý via fino ad arrivare alla soprammenzionata latebra mediante una stratificazione alternata e concentrica. Nell'uovo sodo, il tuorlo bianco rimane sempre pi¨ molle di quello giallo perchÚ vi sono profonde differenze nella loro intima costituzione. Infatti ii secondo consta di granuli molecolari e di sferule microscopiche, molli ed elastiche, mentre il primo di granuli di dimensioni superiori e di sferule pi¨ piccole. Nel nucleo di Pander si accumulano le materie grasse del tuorlo.
The Nucleus of Pander provides the place where the embryo develops. The blastoderm is an early stage of the embryo, present when the egg is laid. The blastoderm is attached to the Nucleus of Pander. The blastoderm grows during the incubation period to ultimately become the chick.
Induction
and improved embryonic development by the nucleus of Pander
in associated avian blastoderm parts: influence of delta or gamma ooplasm.
Callebaut M, Van Nueten E, Harrisson F, Bortier H.
Source
University of Antwerp, Faculty Medicine (Morphology), Anatomy and Embryology,
B-2020 Antwerpen, Belgium. marc.callebaut@ua.ac.be
Abstract
After placing in vitro, central subgerminal ooplasm (containing a central nucleus of Pander) from a quail germ disc of a prelaid egg (before symmetrization) on the upper layer of an isolated chicken antisickle, we observed the induction of a radially oriented preneural plate (without interference of chordamesoblast). This observation suggests the primary existence during the period of symmetrization in utero of an until now unknown temporospatially linked "vertical" effect, emanating from the nucleus of Pander, on the parallel (pre)neural plate anlage forming part of the area centralis in the overlying blastoderm. For comparison, we "sandwiched" in vitro a quail sickle endoblast fragment between the deep side of the upper layer of an isolated chicken antisickle region and a central subgerminal ooplasmic mass. This resulted in a colonization of the subgerminal ooplasmic mass by quail sickle endoblast cells followed by improved neurulation and/or gastrulation phenomena. The latter never occurs in the absence of central subgerminal ooplasm. In both types of experiments there seems to exist a common link between the observed induction phenomena: the presence of delta ooplasm in the involved deep structures. Indeed, the nucleus of Pander contains delta ooplasm as well as the structures derived from it, i.e., endophyll with primordial germ cells and sickle endoblast-derived cells after colonization of the neighboring central ooplasm (present study). Therefore, we think that the preneural plate-inducing effect observed after placing a nucleus of Pander on the antisickle region is due to the presence of a factor in the delta ooplasm that diffuses in the neighborhood. The appearance of gastrulation phenomena in the second type of experiment seems to be due to colonization of the more peripheral part of the central subgerminal ooplasm containing the more superficial and peripheral gamma ooplasm in which Rauber's sickle material can develop. This suggests that the kind of involved ooplasm (delta or gamma) can predetermine the inductive activity of the deep structures that contain it: the central part of the nucleus of Pander and/or endophyll for preneurulation phenomena and sickle endoblast (in the presence of central subgerminal ooplasm) for gastrulation and/or neurulation phenomena.
Journal of Morphology 2004 May;260(2):201-8.
![]()