Vol. 2° - IX.2.
L’endocitosi
Endocitosi č un termine
generale usato per indicare un gruppo di fenomeni legati all’ingresso di
particelle solide o liquide nel citoplasma per invaginazione della membrana.
2.1. Macropinocitosi
Si tratta di un meccanismo
visibile in microscopia ottica mediante il quale la cellula, con un movimento
della membrana, cattura una gocciolina di un liquido. La macropinocitosi
avviene in due modi diversi:
q
per
un movimento di ripiegamento di una lamina ectoplasmatica sul corpo cellulare:
in questo caso si forma un’espansione citoplasmatica stretta, a forma di
lama, che si ripiega sulla gocciolina; segue quindi una fusione con la
membrana plasmatica e la gocciolina viene a trovarsi in una vescicola con
diametro pari a 1÷2 mm
q
per
invaginazione della membrana plasmatica: viene a crearsi un’invaginazione
tubuliforme profonda; le vescicole si formano per strozzature scaglionate
lungo l’invaginazione.
2.2. Micropinocitosi
La membrana si introflette in
una formazione tubuliforme lunga 100 nm con un diametro pari a 30÷50 nm. Il
liquido penetra nel tubulo che, per strozzatura, dŕ origine a microvescicole.
La membrana puň anche introflettersi dando origine direttamente a una
vescicola immersa nel citoplasma, senza che si formi prima il tubulo. In
questo caso si tratta di una microvescicola o vescicola di micropinocitosi.
2.2.a. Le vescicole lisce
Le vescicole lisce corrispondono
a un’endocitosi poco specifica che svolge un ruolo nel trasporto di sostanze
attraverso la cellula. Questo fenomeno si verifica, per esempio, nelle cellule
endoteliali, che sono cellule pavimentose, formanti con la lamina basale la
parete dei capillari.
Le vescicole di micropinocitosi si formano sulla
superficie cellulare in rapporto col lume vasale e quindi col sangue
(superficie luminale) e trasportano, attraverso la cellula, le sostanze
catturate (macromolecole, proteine plasmatiche, anticorpi, micelle
lipoproteiche), liberandole per esocitosi nel mezzo extracellulare che
circonda il capillare.
2.2.b. Gli acantosomi - coated vesicles -
vésicules hérissées
La formazione di coated
vesicles č un processo di internalizzazione specifico di regioni
specializzate della membrana plasmatica associate a un gruppo di recettori,
che si invaginano rapidamente, finendo per isolarsi.
La superficie esterna di tali vescicole č irta di spine
lunghe 15 nm. Questo aspetto č dovuto alla presenza di un reticolo formato
dall’associazione di numerose molecole fibrillari di clatrina (180.000 Da)
che si dispongono alla superficie della vescicola formando esagoni o
pentagoni.
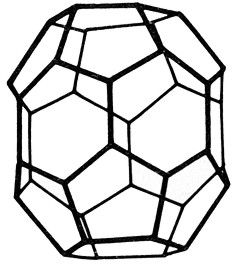
Fig.
IX. 2 - Ricostruzione
tridimensionale di un acantosoma
Il
reticolo esagonale e pentagonale alla superficie della coated
vesicle
č dovuto alla disposizione delle molecole fibrillari di clatrina.
La membrana plasmatica, prima dell’endocitosi, laddove
si formerŕ la coated vesicle, si
ricopre sul versante citoplasmatico di una rete a maglie esagonali e
pentagonali di clatrina. Le zone di depressione appaiono irte di punte e in
queste regioni prendono posto recettori specifici della sostanza destinata ad
entrare nella cellula, che costituisce il ligando.
Il passaggio del ligando nel citoplasma - che costituisce la fase di
internalizzazione, o meglio, di interiorizzazione - č obbligatoriamente
subordinato al legame col suo recettore. Il recettore specifico si
localizzerebbe spontaneamente nella depressione irta di punte, coated
pits, oppure l’accoppiamento recettore-ligando potrebbe realizzarsi in
una zona diversa e l’insieme migrerebbe successivamente verso tale
depressione.
Prendiamo ad esempio il colesterolo. La cellula sintetizza
un recettore specifico per le LDL, low density lipoproteins, sintetizzate nel fegato e circolanti nel
sangue, formate da un core apolare
contenente esteri del colesterolo, circondato da uno strato di fosfolipidi e
proteine. Le LDL si fissano ai recettori di membrana e, dopo 10 minuti di binding,
vengono interiorizzate nelle coated
vesicles che, in seguito alla fusione con lisosomi primari, sono dotate di
attivitŕ enzimatiche. La porzione proteica viene idrolizzata in aminoacidi e
dagli esteri del colesterolo viene recuperato il colesterolo, che potrŕ
servire per la sintesi di strutture membranarie. La sintesi di recettori per
le LDL dipende dal contenuto in colesterolo intracellulare: se questo č
basso, la sintesi di recettori da parte della cellula che necessita di
colesterolo si eleva; si tratta del cosiddetto feedback
o retrocontrollo cellulare.
2.3. La fagocitosi
La fagocitosi č l’insieme dei
fenomeni che portano alla cattura e alla demolizione di particelle solide ed
č stata a lungo considerata come un meccanismo di difesa riservato, nei
vertebrati, a cellule assai specializzate, delle quali rappresenta la funzione
essenziale: i granulociti e i macrofagi, cioč le cellule del sistema
reticoloistiocitario.
Altri tipi di cellule, tuttavia, in certi momenti della
loro vita, per situazioni di necessitŕ - rappresentate per esempio da uno
stato patologico - possono adattarsi a questo sistema di assorbimento di
particelle solide. Questa fagocitosi si esplica non solo verso i batteri, ma
anche per frammenti cellulari.
Quando, per esempio, i batteri riescono a penetrare nell’organismo,
dopo aver superato le molteplici barriere di protezione (epidermide, mucosa
digestiva o respiratoria), vengono immediatamente modificati dagli anticorpi
umorali, le opsonine
[1]
.
L’opsonizzazione, o fissazione di anticorpi specifici
sulla superficie dei batteri, precede obbligatoriamente la loro cattura. I
granulociti migrano nelle zone occupate dai batteri, attirati da una sostanza
chimica di natura ignota; migrano alla velocitŕ di 0,7-1 mm per minuto, fenomeno che č
detto chemiotassi.
I granulociti sono animati da movimenti incessanti:
formazione di pseudopodi, formazione di bolle alla superficie cellulare. Il
contatto precede una fase essenziale, rappresentata dall’adesione, durante
la quale la particella estranea viene immobilizzata contro la membrana
citoplasmatica.
L’ectoplasma invia su tutta la superficie del batterio
dei prolungamenti, che si fondono a limitare una vescicola, il fagosoma,
occupata dal batterio. La membrana del fagosoma prende origine, dunque, dalla
membrana plasmatica. Si tratta della fase reologica. Il fagosoma si trasforma
in fagolisosoma per fusione con lisosomi primari.
La fagocitosi, processo attivo, consuma una quantitŕ
notevole di energia. La spesa energetica necessaria ad incorporare una
particella di polistirolo corrisponde all’energia contenuta in 109 molecole di ATP.
2.4. Inibizione dell’endocitosi
La fagocitosi e la
macropinocitosi sono nettamente rallentate o bloccate da minime concentrazioni
di citocalasina
[2]
,
che annulla le proprietŕ dei microfilamenti e che non č in grado di inibire
la micropinocitosi. Durante l’inibizione i batteri opsonizzati aderiscono ai
macrofagi o ai granulociti, ma non vengono mai interamente circondati dalla
membrana citoplasmatica e restano all’interno di invaginazioni piů o meno
profonde della membrana stessa.
2.5. Ruolo dei microfilamenti nell’endocitosi
La rete di microfilamenti gioca
un ruolo importante nell’internalizzazione svolta dalla membrana. Le
proteine di membrana che aderiscono a una particella o a una cellula batterica
vengono immobilizzate dai filamenti di actina sottomembranari. Il blocco della
pompa del Na+ aumenta la concentrazione intracellulare di Na+
provocando cosě un’onda di depolarizzazione che libera il Ca++
accumulato nei canalicoli del reticolo endoplasmatico.
La presenza di calcio č indispensabile per la
combinazione dell’actina con la miosina: tale combinazione, con formazione
di actomiosina, si accompagna a una contrazione. Poiché questi filamenti sono
fissati sulla faccia interna della membrana, provocano la comparsa di una sua
invaginazione, che si richiude sulle particelle adese alla membrana.
2.6. Il controllo dell’endocitosi
Dal punto di vista quantitativo
il controllo dell’endocitosi č poco conosciuto. Si sa, tuttavia, che certe
sostanze sono capaci di inibire la pinocitosi, mentre altre la stimolano.
Cosě, gli ormoni controllano la pinocitosi delle cellule tiroidee.
2.6.a. Inibizione dell’endocitosi
Inibitori sintetici bloccano l’endocitosi.
Il macrofago risulta incapace di effettuare l’endocitosi quando č mantenuto
in un mezzo con p-fluoro-fenilalanina. L’integritŕ della via glicolitica e
della fosforilazione ossidativa sono indispensabili per l’endocitosi: l’aggiunta
di DNP (dinitrofenolo) la inibisce, ma si tratta di una situazione reversibile
per aggiunta di ATP malgrado sia presente DNP.
2.6.b. Induzione dell’endocitosi
Alcuni coloranti basici e alcune
proteine inducono l’endocitosi. L’induttore č caricato positivamente e si
associa, grazie a forze elettrostatiche, ai mucopolisaccaridi del cell
coat.
L’endocitosi, salvo che per le cellule istiocitarie
(granulociti e macrofagi), avviene in tutte le cellule indipendentemente dall’induzione.
L’endocitosi indotta č infatti un fenomeno tossico o patologico,
esagerazione di un fenomeno fisiologico.
|
sommario |
avanti |
|
|
|