Vol. 2° - XXVIII.9.2.
Tricocromi
Un tempo ritenuti caratteristici componenti dei capelli rosso brillante, i tricocromi sono stati trovati nei peli di parecchi altri mammiferi, compresi i bovini dal pelo rosso, il criceto dorato, il coniglio rosso della Nuova Zelanda e nei cani bastardi rossi.
Per nostra estrema
soddisfazione, possiamo aggiungere che i tricocromi sono presenti anche nelle
piume rosse e fulve del pollo domestico, specialmente nella New Hampshire e nella Rhode
Island red, che costituiscono la fonte migliore per la loro
estrazione e purificazione.
Sono due i tricocromi riscontrati nelle piume rosse e fulve del pollo, uno dei quali è identico al pigmento dei capelli rossi. Dal momento che il colore della piuma non cambia estraendo i tricocromi, il loro ruolo colorante non è ancora del tutto chiaro.
Le piume color salmone del petto della Livorno
dorata e del mantello delle femmine frumento contengono scarsa quantità di
uno dei 2 tricocromi descritti nel pollo; tuttavia sono ambedue assenti nel
mantello brunastro di femmine dotate di genotipo e+
e eb, dorate e perniciate.
Come accade per le feomelanine, esistono parecchi tipi di
peli rossi non contenenti tricocromi: setter irlandese, vitello dell’Ayrshire,
cavia nella varietà completamente rossa. In realtà, sembra che la
distribuzione dei tricocromi nei peli e nelle piume vada di pari passo con
quella delle feomelanine, a causa delle strette relazioni chimiche e
biogenetiche tra le due classi di pigmenti.
Non è ancora certo se i tricocromi siano presenti nella
pelle e nei tessuti extracutanei. Interessante il fatto che essi sono presenti
nell’urina di alcuni pazienti affetti da metastasi di melanoma. I valori più elevati di
tricocromuria sono stati trovati in soggetti con melanoma metastatizzante e
melanosi diffusa con elevato incremento dell’escrezione di cisdopa.
Comunque, non è stato possibile porre alcuna relazione tra il tipo
pigmentario cui appartiene il paziente affetto da melanoma e l’escrezione di
tricocromi.
Non è chiaro se
questi pigmenti siano prodotti dai melanociti o se essi facciano la loro
comparsa come risultato di eventi metabolici che si svolgono in altre cellule,
come i cheratinociti oppure i macrofagi, che possono incorporare i
precursori dei tricocromi, cioè le molecole di cisdopa.
9.2.a. Struttura e proprietà dei tricocromi
L’unità strutturale è
rappresentata da un sistema di anelli di 1,4-benzotiazina che possono esistere
sotto forma di due tautomeri, 2H- e 4H-1,4-benzotiazina. A causa della sua
spiccata instabilità, il composto di origine non è ancora stato isolato,
anche se si è potuto documentare che la forma transeunte predominante è
rappresentata dal tautomero 2H-.
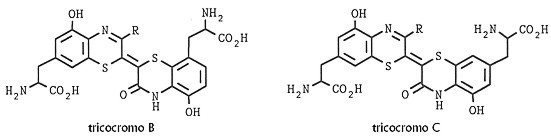
Fig. XXVIII. 5. – Struttura dei tricocromi B e C
I tricocromi più frequenti B e C sono stati isolati per la prima volta dalle piume
rosse delle femmine di New Hampshire e furono successivamente trovati nei
capelli color carota, ma, anche se sono intensamente colorati, rappresentano
solo la frazione minore dei pigmenti feomelanici dei capelli.
I tricocromi E
ed F
apparentemente non sono presenti nei capelli, e non è ancora chiaro se la
piccola quantità trovata nelle piume rosse e nelle altre fonti di pigmento
siano artefatti dovuti all’ossidazione oppure prodotti intermedi di sintesi
che compaiono durante l’isolamento e il trattamento con acidi e alcali.
Da un punto di vista generale i tricocromi sono dotati di
proprietà non comuni, come l’insolubilità
nei solventi organici e nell’acqua, ambedue a pH neutro,
cristallizzazione difficile e forte tendenza all’aggregazione e all’assorbimento.
I tricocromi giallo arancio B e C sono inoltre relativamente instabili in
acidi.
Anche se lo scheletro stereochimico dei tricocromi non è
ancora stato definito, i pigmenti isolati costituiscono verosimilmente una miscela in equilibrio tra due isomeri,
l’isomero trans che predomina al buio e che può
trasformarsi nella forma rossa instabile cis dopo breve irradiazione con luce
solare.
|
sommario |
avanti |
|
|
|