Vol. 2∞ - III.5.
IL NUCLEOLO
Il nucleolo Ë un organulo responsabile della sintesi degli acidi ribonucleici dei ribosomi, presente nel nucleo durante le fasi G1, S e G2. Scompare durante la mitosi.
Il numero dei nucleoli eguaglia di solito il grado di plodia: ci sono 2 nucleoli nelle cellule diploidi, 1 nelle aploidi, 4 nelle tetraploidi e cosÏ via. Spesso perÚ i due nucleoli di una cellula diploide possono fondersi in uníunica struttura, oppure possono esservi numerosi nucleoli per cellula, come accade per le cellule germinali di vari animali.
Negli
oociti degli anfibi, muniti di un nucleo enorme, ci sono migliaia di nucleoli
disposti alla periferia del nucleo, al di sotto dellíinvolucro nucleare.
Al
microscopio ottico il nucleolo si presenta come un granulo molto
rifrangente, rotondeggiate oppure ovalare, situato in posizione eccentrica, in
rapporto con masserelle di eterocromatina (eterocromatina
nucleolo-associata) che lo circondano pi˘ o meno completamente in forma díanello.
Il
nucleolo si presenta con le seguenti strutture:
ÿ cromatina
perinucleolare,
che si dispone a fascia, circondando pi˘ o meno completamente il corpo del
nucleolo. Questa cromatina perinucleolare presenta proprietý fisiche
singolari che permettono di isolare i nucleoli intatti ricorrendo allíultracentrifugazione.
Il calcio indurisce questa zona di cromatina e, in assenza di calcio, il
nucleolo si gonfia e perde le sue caratteristiche morfologiche.
ÿ
corpo
nucleolare,
in genere sferico, che corrisponde alla porzione di nucleolo circondata da
cromatina perinucleolare.
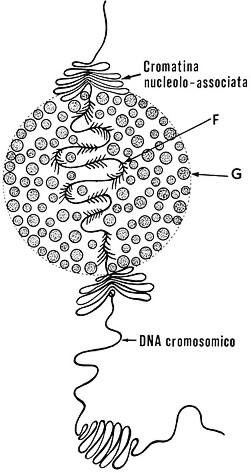
Fig.
III. 22
- Nucleolo in un nucleo intercinetico.
Colorato con azzurro C o con blu di toluidina, il nucleolo si presenta come un granulo denso, omogeneo, con diametro medio di 1-2 mm nelle cellule normali. AllorchÈ in profase si rendono visibili i cromosomi, si puÚ constatare che il nucleolo aderisce a una certa area di un determinato cromosoma.
Al momento della scomparsa del nucleolo, tale area si presenta come un sottile
tratto filamentoso in seno allíeterocromatina del cromosoma: questíarea Ë
detta organizzatore nucleolare,
NOR, in quanto Ë proprio qui che
in telofase si riforma il nucleolo scomparso durante la mitosi. In realtý líorganizzatore
nucleolare corrisponde alla pars fibrillaris
del nucleolo. Talora si dý il nome di organizzatore nucleolare anche allíintero
cromosoma che contiene líarea di cui abbiamo appena parlato.
Líorganizzatore nucleolare spesso si trova nei pressi di una delle estremitý del cromosoma
dove si nota, in tal caso, un sottile tratto filamentoso recante appesa la
parte distale del cromosoma sotto forma di granulo, detto satellite.
Oggi, perÚ, col termine di satellite si indicano sempre
pi˘ frequentemente certe frazioni di DNA altamente ripetitivo che non hanno
nulla a che fare col nucleolo. Nella specie umana le cellule diploidi hanno
ben 10 cromosomi muniti di tale organizzatore; raramente perÚ hanno 10
nucleoli, perchÈ alcuni organizzatori si fondono tra loro, altri invece
restano inattivi non collaborando alla formazione dei nucleoli.
5.1. Biochimica del nucleolo
5.1.a. DNA nucleolare
La replicazione del DNA
nucleolare avviene durante la fase S del ciclo cellulare. La replicazione del
DNA perinucleolare Ë pi˘ precoce e pi˘ rapida rispetto a quella del DNA
intranucleolare.
5.1.b. RNA nucleolari
Nel nucleolo esistono diversi
tipi di RNA che vengono classificati a seconda del loro coefficiente di
sedimentazione espresso in unitý Svedberg. Nei nucleoli delle cellule
epatiche si trovano i seguenti RNA: RNA 45S, 35S, 28S, da 8 a 16S, da 4 a 7S,
infine il 18S, isolato dal nucleoplasma. I differenti RNA corrispondono a
tappe maturative.
5.1.c. Proteine nucleolari
Il nucleolo contiene due tipi di
molecole proteiche: proteine acide e istoni. Le loro funzioni sono poco
conosciute.
5.1.d. Enzimi nucleolari
Nel nucleolo esistono enzimi che
intervengono in importanti processi biochimici di sintesi e maturazione dellíRNA.
5.2. Ruolo del nucleolo
5.2.a. Trascrizione dellíRNA ribosomiale Û rRNA
Ricorrendo a esperimenti di citofisiologia comparata diventa talora facile comprendere un meccanismo biologico di interesse generale.
Tra i vari tipi di Xenopus laevis,
un rospo africano, ne esiste uno che possiede un solo nucleolo, mentre
abitualmente lo Xenopus laevis
possiede due nucleoli per nucleo. La presenza di un solo nucleolo Ë una
caratteristica ereditaria: se si incrociano due rospi con cellule dotate di un
solo nucleolo, il 25% della progenie di girini possiede nuclei con due
nucleoli, il 50% nuclei con un nucleolo e il 25% nuclei senza alcun nucleolo.
Líultimo gruppo muore dopo quattro giorni dalla nascita,
ma la vita dei morituri Ë lunga quanto basta per realizzare il seguente
esperimento: si pongono i tre tipi di girino in un mezzo contenente precursori
radioattivi dellíRNA. I girini con 1 o 2 nucleoli incorporano i precursori e
sintetizzano RNA messaggero, RNA transfer e RNA ribosomiale; quelli con nuclei
sprovvisti di nucleolo sintetizzano solo RNA messaggero e RNA transfer. Quindi
i girini senza nucleolo hanno perso la possibilitý di sintetizzare rRNA. Le
zone fibrillari sono la sede in cui avviene la sintesi di RNA, e ciÚ Ë in
perfetto accordo con líosservazione che tali zone sono strettamente
giustapposte al materiale cromosomico.
5.2.b. La maturazione dellírRNA
Il DNA nucleolare che serve da
stampo per la sintesi dellírRNA
Ë una porzione di DNA del cromosoma che si trova nellíorganizzatore
nucleolare,
a livello della porzione fibrillare. Il nome di rDNA, o DNA ribosomiale, spetta ai geni
responsabili della sintesi di rRNA corrispondente allírRNA 45S.
Questo precursore, in seguito a maturazione, dý origine a
una molecola di RNA 18S e a uníaltra di 28S, entrambe metilate e associate a
proteine ribosomiali. Tali particelle
di RNP
ribosomiali (40S per líRNA 18S, 65S per líRNA 28S) passano nel citoplasma
attraverso i pori nucleari per associarsi e formare i ribosomi.
5.2.c. Regolazione della trascrizione dellírRNA
La trascrizione di rRNA dipende dalla quantitý di ribosomi presenti nel
citoplasma. La tioacetamide (CH3
CSH3) stimola la sintesi di rRNA, che risulta 10 volte
superiore a quella di cellule non trattate; provoca inoltre lesioni
lisosomiali con liberazione progressiva di enzimi, quali proteasi e
ribonucleasi, portando a una distruzione di ribosomi che diminuiscono del
50-70% e conseguente ipertrofia del nucleolo da aumentata produzione di rRNA.
La regolazione dellíattivitý nucleolare si basa sulla seguente ipotesi:
esisterebbe un sistema di retroazione, o feedback,
per cui un prodotto ribosomiale o gli stessi ribosomi potrebbero funzionare da
repressori genici con sblocco dellíinibizione in seguito alla loro
distruzione.
5.2.d. Nucleolo e mRNA
Un nucleolo funzionante Ë necessario per il trasferimento dellímRNA dal
nucleo al citoplasma.
Líinattivazione del nucleolo con un microfascio di luce UV inibisce il
passaggio di mRNA in seno al citoplasma.
5.2.e. Preparazione alla mitosi
Il nucleolo Ë necessario per un
corretto svolgersi della mitosi. In effetti, importanti alterazioni dei nucleoli provocano il blocco delle
cellule in fase G2, che precede la mitosi o fase M. Il trattamento
del nucleolo con un microfascio di luce UV inibisce la divisione cellulare. Se
durante líinterfase e subito prima della scomparsa del nucleolo la sintesi
proteica Ë bloccata con acticromo o con cicloesimide, la mitosi ha luogo, ma
i nuclei non possono riformarsi.
5.3. Effetti delle variazioni del nucleolo nel Pollo
Le variazioni a carico delle
dimensioni del nucleolo sono comuni sia nellíuomo che negli altri animali.
Nonostante ciÚ, non Ë ancora chiaro il significato nonchÈ la causa di tali
variazioni. Le copie di geni che nei vertebrati codificano gli RNA
ribosomiali, o rRNA 18S, 5.8S e 28S, sono numerose e sono raggruppate in
corrispondenza di parecchi siti cromosomici, noti come regioni
dellíorganizzatore nucleolare o NOR.
I ribosomi vengono prodotti in questi siti in seguito alla
trascrizione dellírDNA, o DNA ribosomiale, sotto forma di rRNA e successiva
formazione delle subunitý preribosomiali. I geni per lírRNA sono presenti in numero molto
elevato: circa 400 nella Drosofila, nel pollo e nellíuomo, 1.000 negli
anfibi, 10.000 nelle piante. Questo numero elevato garantisce il meccanismo
dellíabbondante produzione di rRNA indispensabile alla sintesi proteica e
alla divisione cellulare, ed Ë una caratteristica costante di gran parte dei
genomi. Proprio a causa della funzione strutturale e catalitica svolta durante
la sintesi proteica, gli rRNA suddetti sono essenziali per la
differenziazione, lo sviluppo e la crescita.
Anche se Ë noto il numero di geni codificanti per lírRNA,
non Ë ben chiaro cosa possa accadere allo sviluppo e alla vitalitý dei
vertebrati omeotermi quando si verificano delle variazioni, specialmente in
caso di riduzione numerica dei geni codificanti. A questo scopo Delany e coll.
(1994) hanno studiato il potenziale di sviluppo in embrioni di pollo
caratterizzati da ben definite deficienze numeriche a carico dei geni per gli
rRNA. I risultati sono stati ottenuti impiegando un ceppo caratterizzato da un
polimorfismo a carico delle dimensioni del nucleolo, che riflette a sua volta
il polimorfismo delle dimensioni dei raggruppamenti di rDNA.
Gli embrioni caratterizzati da un nucleolo grande e da un
nucleolo molto piccolo (Pp) sono eterozigoti per un raggruppamento ridotto di
rDNA (+/p1) e contengono circa il 66% dellíequivalente per i geni
dellírRNA degli individui normali (+/+), i quali sono dotati di due nucleoli
di dimensioni normali e identiche (PP). Gli embrioni +/p1
si
sviluppano e crescono in modo regolare.
Gli
embrioni con due nucleoli molto piccoli (pp) sono omozigoti per la
deficienza di rDNA (p1/p1) e contengono circa il 45%
dellírDNA. Questi embrioni si
arrestano in fasi
molto precoci di sviluppo, e precisamente in fase di gastrula. Le ricerche hanno anche
dimostrato che per raggiungere il successivo stadio di blastula sono necessari
e cruciali adeguati livelli di rRNA. Gli eterozigoti Pp riescono a formare
quantitý adeguate di rRNA. Nel Gallus
domesticus i raggruppamenti di rDNA
hanno una dimensione e uníorganizzazione molecolare simile a quella dei
mammiferi, e tutti i geni per lírRNA risiedono su un singolo paio di
cromosomi. Nel pollo i loci dellíorganizzatore
nucleolare occupano circa 6.000 kb del
braccio lungo del microcromosoma 17. Líaltro braccio di questo cromosoma Ë
occupato dal Complesso Maggiore di Istocompatibilitý o locus B. I loci dellíorganizzatore
e il locus B distano circa 10 kb.
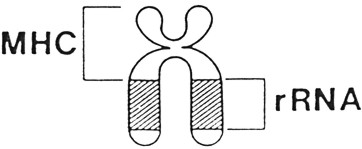
Fig.
III.
23
- Microcromosoma 17 del pollo.
I
loci dellíorganizzatore nucleolare, o NOR, occupano circa 6.000
kb del braccio lungo.
|
sommario |
avanti |
|
|
|