Vol. 2░ - XIX.11.1.
Emoglobina e mioglobina
Lĺevoluzione dellĺemoglobina
e della mioglobina Ŕ un esempio classico in cui si pensa che essa si sia
verificata attraverso un processo rappresentato da successive duplicazioni del gene. Ambedue queste molecole proteiche sono importanti per il
trasporto dellĺossigeno nei vertebrati: lĺemoglobina, contenuta nei
globuli rossi nella quantitÓ di 280 milioni di molecole per eritrocita,
assicura il trasporto dellĺossigeno dai polmoni ai tessuti; la mioglobina
rappresenta una riserva di ossigeno prevalentemente per il muscolo
scheletrico.
Nellĺuomo,
la forma predominate di emoglobina
dellĺadulto Ŕ rappresentata da due coppie di catene polipeptidiche,
a e b globina, costituite ognuna da
due subunitÓ di catene polipeptidiche uguali (quindi a1
e a2
- b1
e b2),
ma esistono anche delle piccole quantitÓ di varianti dellĺemoglobina
normale, che finora assommano a 90. Altre forme di emoglobina sono presenti
nel feto (forma g) e nellĺembrione durate le
primissime fasi di sviluppo (forma e).
Nelle specie di mammiferi e di uccelli in cui finora lĺemoglobina
Ŕ stata esaminata, Ŕ possibile assegnare le catene di globina a uno dei due
tipi base, a
e b. La suddivisione si basa sul
grado di omologia tra la globina in questione e il tipo a oppure b, e si pensa che tale omologia
rifletta la loro origine evoluzionistica.
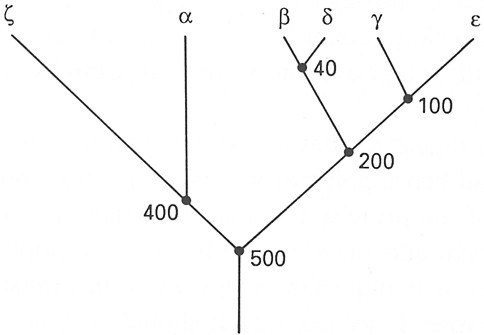
Fig.
XIX. 9 ľ
Probabili relazioni evoluzionistiche delle globine umane a
b,
g,
d,
e
e z
I numeri indicano i milioni di anni dallĺinizio della divergenza.
Si Ŕ dellĺavviso
che a ogni punto delle ramificazioni si sia verificata una duplicazione
genica.
Nellĺuomo il tipo
b
comprende le globine b,
g, d ed e, mentre il tipo a
comprende le globine a
e z. Si pensa che le globine
appartenenti al tipo a
siano andate incontro a divergenza 400 milioni dĺanni fa, prima del distacco
di mammiferi e uccelli dai rettili, mentre le globine del tipo b hanno subito lo stesso processo
di divergenza circa 200 milioni dĺanni fa, contemporaneamente al distacco
filogenetico dei mammiferi e degli uccelli.
Nel
pollo domestico esistono due forme principali di emoglobina
adulta, A e D, HbA
70% e HbD 30%, nonchÚ due forme minori, H
e L, HbH
- HbL presenti in tracce. Come nel
caso dei mammiferi, esistono almeno 3 forme di emoglobina embrionale, presenti
nei globuli rossi che si sviluppano nel sacco vitellino fino al 5░ giorno dĺincubazione.
Tra il 5░ e il 12░ giorno le cellule embrionali della serie rossa vengono
gradualmente sostituite da eritrociti normali contenenti emoglobina adulta.
Recentemente sono stati scoperti altri tipi di globine che appartengono al
tipo a
oppure al tipo b e queste catene vengono
codificate da due
famiglie multigeniche.
Una famiglia multigenica consiste in un gruppo di geni tra loro correlati, e
raggruppati tra loro su particolari cromosomi. Si pensa che questi geni siano
derivati da successive
duplicazioni geniche verificatesi durante la meiosi a causa di un
crossingover ineguale. Un buon numero di famiglie multigeniche Ŕ stato
identificato, e i geni delle globine appartenenti al tipo b sono stati mappati usando una
banca genomica
[1]
del pollo domestico.
Le famiglie geniche per la globina sono spesso considerate
come un paradigma nello studio dellĺespressione
genica regolata dallo sviluppo dellĺorganismo.
I globuli rossi degli uccelli sono particolarmente utili sotto questo profilo
in quanto presentano degli stadi di sviluppo ben definiti nella loro
successione. Dal momento che anche il tipo a
della globina Ŕ stato sottoposto a sequenziamento, Ŕ possibile studiare in
modo dettagliato la regolazione dipendente dallo sviluppo.
Lĺespressione genica dipende in primo luogo dalla trascrizione, e questo fatto richiede lĺunione della RNA polimerasi ai siti promotori adiacenti al gene. In questo punto esistono anche delle sequenze di DNA poste al di fuori della regione del promotore, note come enhancers, in grado di incrementare la velocitÓ di trascrizione, e sono spesso molto importanti nella regolazione genica durante il processo di sviluppo dellĺorganismo, e possono essere specifici a seconda dei tipi di tessuto. Sono in grado di agire a distanza, cioŔ non hanno bisogno di essere adiacenti al gene, e possono essere localizzati dal lato 5ĺ oppure 3ĺ del gene stesso.
Gli enhancers interagiscono con proteine specifiche, e si pensa che in
qualche modo rendano il gene pi¨ accessibile alla RNA polimerasi. Si Ŕ visto
che gli enhancers sono adiacenti ad
ambedue le famiglie per a
e b globina, localizzate su
cromosomi differenti: sul cromosoma 1 o 2 la famiglia a, su un piccolo macrocromosoma o
forse su un microsoma grande la famiglia b.
Inoltre, considerando lĺevoluzione di un particolare gruppo di proteine in
seno a una specie, Ŕ possibile tessere delle relazioni tra gli stadi
evoluzionistici in seno alla specie nei confronti dellĺevoluzione e della
divergenza di specie, di generi e di famiglie fra loro.
Si pensa che la catena di e
globina si sia evoluta dalla b
globina per duplicazione genica. In base alle differenze tra gli aminoacidi di
queste globine, il gene della e
globina del pollo domestico si Ŕ evoluto sin dal momento dellĺorigine della
classe degli uccelli, e nei mammiferi le globine di tipo e si sono evolute dal loro tipo b in modo indipendente.
Il
pollo domestico Ŕ alquanto insolito per il fatto di possedere due tipi di globine
a,
aA
e aD,
ciascuna presente in discreta quantitÓ. La globina aD
Ŕ presente nel nand¨, nello struzzo, nel fagiano, nel passero, nellĺoca
selvatica e nel condor delle Ande, mentre altre specie di uccelli, come
pinguino, egretta, piccione, cuculo e parrocchetti non posseggono la globina aD,
assente anche nei mammiferi. Si pensa che gli uccelli che non presentano la
globina aD
hanno perso la capacitÓ di esprimere il relativo gene, espresso precocemente
nei loro antenati. Non Ŕ certo se questa emoglobina sia essenziale nel pollo
domestico. Sta di fatto che negli ultimi stadi embrionali lĺemoglobina aD
Ŕ presente in proporzioni pi¨ elevate rispetto allĺadulto e che la
presenza dellĺemoglobina aD
aumenta lĺaffinitÓ del sangue per lĺossigeno.
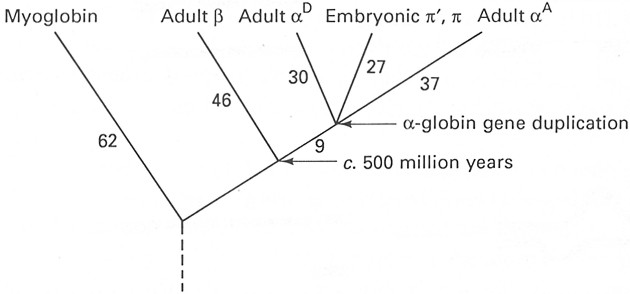
Fig. XIX. 10 ľ Evoluzione delle globine - mioglobina ed emoglobina - nel pollo domestico.
Nonostante la mioglobina sia stata sottoposta a sequenziamento in molte specie di vertebrati, Ŕ stata studiata secondo lo stesso principio solo in due specie di uccelli: pollo domestico e Pinguino imperatore, Aptenodytes forsteri. Pertanto al momento attuale non abbiamo informazioni sufficienti, basate su questa sostanza, allo scopo di chiarire lĺevoluzione in seno alla classe degli uccelli. Nel caso delle emoglobine Ŕ possibile non solo paragonare le sequenze aminoacidiche, ma anche le sequenze dei nucleotidi, dal momento che i dati sulle sequenze sono disponibili da un elevato numero di globine a e b, come pure da un certo numero di pseudogŔni [2] della globina.
Cosý diventa possibile paragonare il numero di cambiamenti a
carico della sequenza nucleotidica, che si manifesta in un cambiamento nella
sequenza degli aminoacidi, con il numero di mutazioni che non provocano alcun
cambiamento, che sono le cosiddette sostituzioni
silenti. I dati a disposizione depongono per una prevalenza di mutazioni
nella sequenza nucleotidica con mutazione nella sequenza aminoacidica rispetto
alle sostituzioni silenti. Tutto
ci˛ conforta la teoria secondo cui lĺevoluzione della globina Ŕ il
risultato di una selezione naturale che si Ŕ svolta su mutazioni di geni
duplicati, e si contrappone alla visione neutralista secondo cui una deriva
genetica casuale sarebbe responsabile del cambiamento nella sequenza
aminoacidica.
|
sommario |
avanti |
|
|
|