vol. 3° - VIII.
UOVO
E
APPARATO GENITALE FEMMINILE
|
|
Nella maggioranza degli uccelli si sviluppano solo l’ovaio
e l’ovidutto di sinistra, mentre quelli di destra sono presenti allo stato
vestigiale. Perň l’ovaio di destra puň essere stimolato ad accrescersi
sino a trasformarsi in un organo maturo se l’altra gonade viene asportata
dopo il 30°
giorno di vita. Se nel pollo l’ovariectomia sinistra č praticata piů
precocemente, essa č seguita dalla formazione di un ovotestis a destra, che
in alcuni casi puň presentarsi come un testicolo funzionante.
1. OVAIO SINISTRO
L’ovaio di sinistra raggiunge
il massimo sviluppo quando la gallina si trova in attivitŕ riproduttiva.
Presenta forma irregolare, simile a quella di un grappolo d'uva: gli acini
sono rappresentati dai follicoli oofori i quali si staccano in
modo piů o meno distinto dalla superficie dell'organo. I follicoli piů
grandi sono ben rilevati e sono uniti al resto dell'ovaio mediante il
peduncolo. Nella gallina in ovulazione sono costantemente presenti da 4
a
6
follicoli
di diametro superiore a 30 mm, che da soli sono sufficienti
a determinare l'aumento del peso ovarico durante il periodo riproduttivo.
L’ovaio č contenuto nella cavitŕ addominale, sospeso
alla volta di tale cavitŕ attraverso un corto mesovario, doppia lamina
peritoneale nel cui contesto sono presenti anche fibrocellule muscolari lisce.
Dorsalmente contrae rapporti con il rene sinistro e con parte del destro, con
l’aorta e con la vena cava. Ventralmente č contornato dal sacco aereo
addominale sinistro che, portandosi verso destra, va a saldarsi al mesovario,
sicché la gonade viene a essere contenuta nella tasca ovarica.
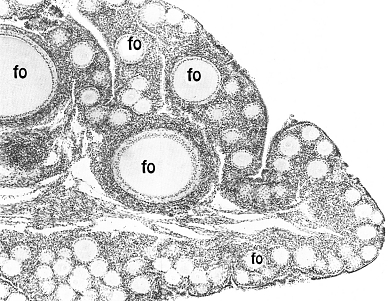
Fig.
VIII. 1 - Ovaio di gallina a piccolo ingrandimento.
fo = follicolo ooforo
Lo sviluppo dell'ovaio diventa significativo al
raggiungimento della maturitŕ sessuale che in genere si verifica dopo 18-20
settimane
dalla nascita. Inizialmente l’accrescimento dell'organo č relativamente
lento, per ricevere una rapida spinta all’avvicinarsi della maturitŕ quando
il suo peso sarŕ di 40-60 g.
A occhio nudo in un ovaio maturo si possono contare 2.500
follicoli
di varie dimensioni; ma l’osservazione microscopica ne indica oltre 12.000
e calcoli teorici depongono per valori che oscillano intorno al milione.
1.1. Struttura dell'ovaio
L'ovaio č costituito
essenzialmente da due distinte porzioni:
·
corticale,
che si trova in superficie e che accoglie i follicoli oofori in vario stadio
di maturazione
·
midollare,
che č profonda e in cui abbondano i vasi sanguigni.
La superficie della gonade č rivestita dall’epitelio
germinativo, tra le cui cellule sono accolti numerosissimi oogoni in attesa
dei processi maturativi.
I follicoli oofori hanno parete piuttosto
complessa costituita da piů strati che si rendono ben manifesti durante la
maturazione: sino a quando sono di piccole dimensioni rimangono nello spessore
della corticale e man mano che la maturazione prosegue vanno sempre piů
sollevandosi sulla superficie dell'organo, cui restano sempre vincolati dal
peduncolo. Per la presenza del peduncolo, negli uccelli le caratteristiche del
follicolo sono del tutto particolari.
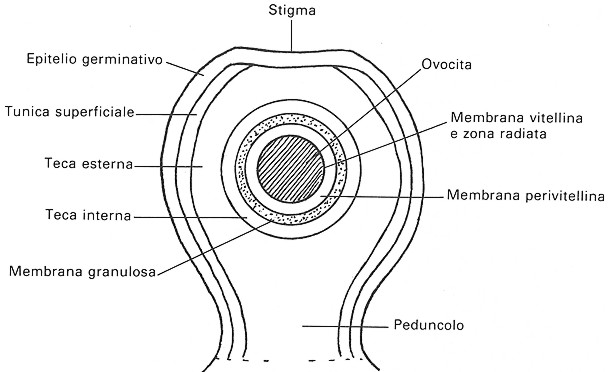
Fig. VIII. 2 - Struttura del follicolo ooforo
La parete di un follicolo prossimo all'ovulazione
appare costituita dai seguenti strati, che cosě si susseguono a partire dallo
strato piů profondo:
·
membrana
vitellina: deriva dalla membrana plasmatica dell’oocita e si
presenta come una rete grossolana di fibre; apparentemente, dopo la
fecondazione subisce scarse modificazioni
·
zona
radiata: si rende evidente nei follicoli che hanno raggiunto un
diametro superiore a 7 mm; ha uno spessore di 5
mm ed č composta da pliche della
membrana vitellina in cui si immette il citoplasma oocitario; proiezioni della
membrana perivitellina si spingono tra queste pliche e possono anche essere
assunte dall’oocita
·
membrana
perivitellina: č una sottile zona acellulare, probabilmente secreta
dalla soprastante membrana granulosa; ha una struttura amorfa, anche se puň
contenere dei corpuscoli in prossimitŕ della zona radiata; si fa piů
evidente nei follicoli prossimi all'ovulazione
·
membrana
granulosa: č composta da elementi cellulari che nei follicoli
molto piccoli o in quelli maturi sono disposti in un solo strato, mentre ne
presenta piů strati nei follicoli di medie dimensioni; all'esterno poggia su
una spessa e ben evidente membrana basale amorfa
·
teca
interna: complessa č la sua costituzione; la parte piů
profonda, contigua alla membrana basale amorfa, č composta da connettivo
lasso con prevalenza di fibre collagene; nella regione intermedia abbondano i
fibroblasti, mentre in quella esterna si trovano numerose cellule a citoplasma
vacuolizzato - le cellule luteiniche - ricche di lipidi e con funzione
endocrina
·
teca
esterna: č costituita da connettivo lasso che comprende molti
fibroblasti forniti di granuli posti a ridosso della membrana cellulare
·
tunica
superficiale: č fornita dallo stroma ovarico; si tratta di un
connettivo lasso molto vascolarizzato
·
epitelio
germinativo: si trova applicato alla superficie esterna del follicolo e talora puň mancare.
Sulla superficie libera dei follicoli di diametro
maggiore a 4 mm si distingue una regione piů chiara, povera di vasi: lo stigma.
Č questo il punto in cui il follicolo si rompe al momento dell'ovulazione in
modo da permettere la deiscenza dell'uovo. Dopo l'ovulazione le pareti del
follicolo collabiscono e nonostante non diano origine a un corpo luteo come
nei mammiferi esse si mantengono attive per un certo tempo secernendo ormoni
steroidei. Queste strutture sono indicate come follicoli
postovulatori e regrediscono rapidamente in circa 6 giorni.
Non tutti i follicoli giungono a maturazione: una quota
notevole, in differenti stadi di sviluppo, degenera e viene riassorbita. Si
tratta dei follicoli atresici, i quali danno luogo a
peculiari quadri morfologici che variano a seconda delle dimensioni del
follicolo colpito dall'atresia. In quelli piů grandi spesso il vitello
fluisce fra le teche, dove viene riassorbito.
Nello stroma ovarico - sia nella porzione corticale che
midollare - si trovano gruppi di cellule
interstiziali
a funzione endocrina. Questi elementi possono avere origine dalle cellule
luteiniche della teca interna dei follicoli postovulatori e atresici, oppure
direttamente dai cordoni sessuali secondari della gonade embrionale.
1.2. Oogenesi
I compiti dell'ovaio consistono
nella produzione delle cellule germinali femminili e in una complicata
attivitŕ endocrina da cui dipendono funzioni molto importanti, quali la
vitellogenesi e la regolazione dello sviluppo dell'ovidutto.
Nella formazione e nella maturazione dei gameti femminili
si distinguono tre fasi successive: moltiplicazione, accrescimento,
maturazione.
La moltiplicazione interessa gli oogoni il cui
numero aumenta per normale mitosi, probabilmente limitata al solo periodo
embrionale. Quando quest’attivitŕ č terminata, gli oogoni sono pronti alle
fasi successive e vengono indicati come oociti.
L'accrescimento si identifica con la deposizione
nel citoplasma di un'enorme quantitŕ di materiale di riserva - il vitello - che
sarŕ utilizzato dall'embrione. L'accrescimento comprende tre fasi:
·
un
periodo di mesi o anni in cui i fenomeni procedono con estrema lentezza e
portano alla deposizione soprattutto di grassi neutri
·
una
fase intermedia di circa 60 giorni con accumulo di proteine
·
un
periodo terminale di appena 7-11 giorni, culminante con l’ovulazione,
durante il quale viene aggiunta al citoplasma la massa del vitello molto ricco
di grassi, per cui in pochi giorni l’oocita passa da 0,5
g
a 19
g
di peso.
Gli oociti che iniziano l’accrescimento presentano nel
citoplasma, a ridosso del nucleo, il corpo di Balbiani,
costituito da citomembrane, strutture di Golgi, vescicole di varie dimensioni
e mitocondri. Nella matrice citoplasmatica abbondano fini granuli, gocciole
lipidiche e strutture filamentose.
Negli oociti che hanno raggiunto 0,3
mm
di diametro il corpo di Balbiani si disperde nel citoplasma e i suoi
componenti vanno ad applicarsi all'interno della membrana cellulare. In tale
zona si trovano anche vescicole di Golgi, mitocondri, gocciole lipidiche e
alcuni organuli caratteristici indicati come lining bodies
[1]
e fusi vitellini. Questi ultimi, con le gocciole
lipidiche, rappresentano il primo vitello.
Durante la seconda fase di sviluppo dell’oocita - da 2
a
6
mm
di diametro - comincia la sintesi del vitello vero.
Il nucleo dell’oocita assume posizione eccentrica e nel citoplasma si
formano numerosi vacuoli forniti di membrana. In alcuni di questi inizia la
deposizione di granuli e di gocciole osmiofile
[2]
,
molto ricche in proteine: si tratta del vitello bianco.
Dal citoplasma scompaiono le gocciole lipidiche e compaiono dei granuli, forse
di glicogeno, concentrati sotto la membrana cellulare.
Negli oociti ad accrescimento rapido l’accumulo di
vitello diventa massivo e si interrompe probabilmente solo con l’ovulazione.
Gran parte del materiale destinato a essere deposto nel citoplasma proviene
dal fegato e penetra nell’oocita con modalitŕ non ancora precisate. Secondo
alcuni esso si limita a diffondere dai vasi tecali per giungere passivamente
alla cellula uovo; secondo altri quest'ultima interverrebbe attivamente
attraverso fenomeni di pinocitosi.
Nella cellula matura il vitello o tuorlo
occupa il citoplasma e appare composto di grosse sferule del diametro di 25-150
mm distribuite in una fase
continua. Sia in queste che nella fase continua sono contenuti numerosissimi
granuli piů piccoli dotati di un diametro di 2 mm. Il meccanismo di formazione
di questo vitello, indicato come vitello giallo,
non č ben noto. Č stato suggerito che le sferule derivino dalla progressiva
trasformazione delle vescicole citoplasmatiche circondate da membrana. Le
tappe iniziali del processo richiedono la vicinanza dei mitocondri che perň
successivamente vanno a concentrarsi nel polo animale dell’oocita.
Con la vitellogenesi si
verificano modificazioni nella composizione chimica del vitello: dapprima č
ricco di grassi neutri, quindi di proteine nella fase di vitello bianco,
infine, quando diventa giallo, contiene soprattutto grassi.
|
Stadi
di sviluppo dell'oocita e della deposizione del vitello |
|||
|
Stadio |
Diametro
in mm |
Aspetto citologico |
Tipo di vitello |
|
1 |
0,05¸0,2 |
Corpo di Balbiani |
Principalmente
gocciole lipidiche di grassi neutri |
|
|
0,2¸0,3 |
Corpo di Balbiani disperso |
|
|
|
0,3¸1 |
Lining bodies e fusi vitellini |
|
|
2 |
2¸3 |
Vacuoli citoplasmatici |
Deposizione
principalmente di proteine, ma anche di alcuni lipidi (vitello bianco) |
|
3 |
3¸6 |
Sfere di vitello |
|
|
4 |
6¸9 |
Inizia la formazione del vitello vero |
Grande
quantitŕ di lipidi (vitello giallo) |
|
|
9¸35 |
Rapido accumulo del vitello |
|
|
da
Botte & Pelagalli |
|||
Durante la maturazione
propriamente detta si giunge alla formazione del pronucleo femminile aploide.
Le tappe comprendono la meiosi 1Ş e la
meiosi 2Ş.
La prima divisione meiotica consiste nella riduzione del numero cromosomico e
conduce a un patrimonio aploide a partire da una costituzione diploide, detta
perciň divisione
riduzionale, mentre la meiosi seconda determina la separazione dei
cromatidi e rappresenta la divisione equazionale.
La divisione riduzionale, dalla quale prendono origine un
oocita secondario e un globulo polare, avviene 2 ore
prima dell'ovulazione, anche se i fenomeni che portano a essa sono giŕ
individuabili 24 ore prima. Questa fase della meiosi sarebbe
controllata dall'ormone luteinizzante, LH.
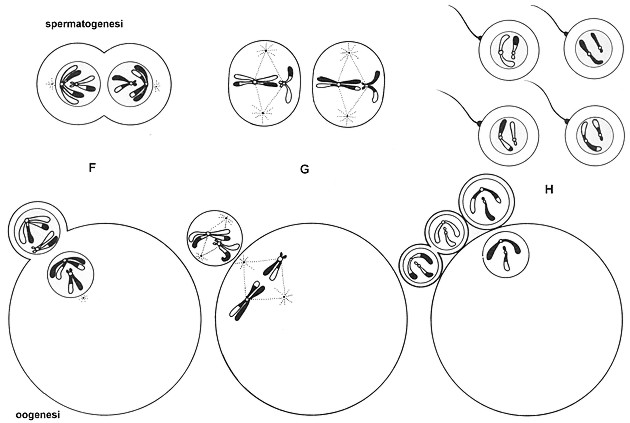
Fig.
VIII. 3 - Oogenesi e spermatogenesi.
F
prodotti della prima divisione meiotica che stanno separandosi
G
prodotti
della seconda divisione meiotica
H
risultato finale della meiosi:
nella femmina si ottengono un uovo e 3
corpuscoli polari,
mentre il maschio produce quattro spermatozoi.
La divisione equazionale si
verifica nell'ovidutto, quindi dopo l’ovulazione, e sembra richiedere l’intervento
dello spermatozoo. Alla fine della divisione equazionale si ha l’espulsione
di un secondo globulo polare e la formazione di un pronucleo femminile che č
pronto a fondersi con il nucleo dello spermatozoo.
1.2.a. Composizione del vitello giallo
Il vitello giallo č composto
per il 50% da acqua; il rimanente 50%
č
rappresentato da lipidi e proteine in proporzione 2:1
che rappresentano i solidi. Nei granuli č contenuto il 23%
dei
solidi: una fosfoproteina, la fosvitina,
e due lipoproteine ad alta densitŕ, le lipovitelline.
A queste ultime č legata gran parte degli ioni calcio e ferro.
Se il tuorlo viene centrifugato, nel sopranatante troviamo oltre a
gran parte dell'acqua un complesso di proteine tra le quali
predomina una lipoproteina a bassa densitŕ -
LDF = low density fraction -
che rappresenta il 65% dei solidi del sopranatante e il 95%
dei
lipidi. Il 10% delle proteine č costituito dalle levitine,
che hanno strette somiglianze con alcune proteine del sangue (globuline,
glicoproteine e albumina). Tralasciamo la dettagliata composizione del tuorlo,
compito dei testi di fisiologia. Dobbiamo invece accennare al contenuto in colesterolo
che č pressoché costante nelle uova deposte da ogni gallina, oscillando da 4
a
10
mg/g
di uovo, potendo variare a seconda delle razze e del ceppo. Le uova a guscio
bianco hanno un contenuto in colesterolo minore rispetto a quelle a guscio
marrone, mentre il tasso maggiore č raggiunto nell’uovo di Araucana.
Sul contenuto in colesterolo puň influire il ritmo della
deposizione, per cui le buone ovaiole danno uova con meno colesterolo. Altro
fattore importante č l’alimentazione, ma di questo parleremo in seguito a
proposito della selezione di linee caratterizzate da uova a basso tenore di
colesterolo.
1.2.b. Controllo della sintesi del vitello
Gran parte dei componenti del
vitello giallo č sintetizzata dal fegato
sotto l’influsso degli steroidi sessuali, essenzialmente estrogeni.
Il fegato riversa i componenti nel sangue e di qui vengono trasferiti
all'oocita in accrescimento.
Nelle cellule epatiche sono state recentemente individuate
modificazioni morfologiche e biochimiche certamente connesse alla
vitellogenesi. Durante questo periodo il contenuto in grassi raddoppia e tutte
le strutture legate alla sintesi proteica si fanno piů evidenti. Inoltre nel
plasma sono state messe in evidenza delle complesse fosfolipoproteine che
per composizione richiamano l’LDF e la fosvitina del tuorlo.
1.3. Ovulazione
Negli
uccelli lo sviluppo postnatale delle gonadi e la loro attivitŕ nella vita
adulta sono controllati dall'apparato endocrino attraverso l’asse
ipotalamo-ipofisario,
sul quale agisce un complicato insieme di fattori esogeni. Anche per il pollo
si verifica qualcosa di analogo a quanto accade all’uomo e che č
stigmatizzato nei Carmina Burana. Infatti il testo medievale, basato
sull’esperienza e del tutto ignaro del gioco ormonale, č piuttosto
categorico:
Tempore
brumali, vir patiens,
Animo vernali, lasciviens.
Latino facile facile: d’inverno, quando c’č la bruma,
l’uomo se ne sta quieto, ma quando la primavera incalza si dŕ alla pazza
gioia. In modo non diverso si comportano i nostri riproduttori.
Pare che negli uccelli, come nei mammiferi, la rottura del
follicolo, seguita dall’ovulazione, si verifichi secondo due modalitŕ:
provocata e spontanea.
1.3.a. Ovulazione provocata
L’ovulazione provocata č
presente nella femmina di piccione, che depone due uova con l’intervallo di
un giorno e con una cadenza ciclica della durata di 35-40 giorni.
Questa deposizione naturale necessita di un evento esterno: coito, eccitamento
visivo. Se alla femmina di piccione vengono sottratte le uova cosě deposte -
rispettando cioč modalitŕ e sequenza - dopo 8 giorni deporrŕ altre 2 uova e
continuerŕ con la stessa cadenza ravvicinata se esse vengono sistematicamente
rimosse dal nido. Possiamo pertanto concludere che entrano senz’altro in
gioco fattori nervosi attivi sull’ipofisi anteriore, e possiamo anche
arguire che l’attivitŕ ipofisaria spontanea non sarebbe in grado di
comandare un’ovodeposizione ravvicinata.
1.3.b. Ovulazione spontanea
Questo tipo di ovulazione č
caratteristico della gallina, la cui attivitŕ preipofisaria spontanea č in
grado di regolare l’ovodeposizione senza intervento di stimoli esterni.
Bisogna aggiungere che, sia nella gallina, sia in altri animali da cortile a
deposizione spontanea come l’anatra, certi fattori esterni, in particolare la
luce,
sono in grado di accrescere in modo notevole l’attivitŕ ipofisaria e di
anticipare la deposizione in femmine prepuberi, o di far deporre le femmine
adulte al di fuori della normale stagione sessuale. La deposizione puň essere
stimolata con
l’alimentazione: proteine animali e vegetali, alcuni sali, olio di
fegato di merluzzo![]() , da non scordare il cibo sminuzzato che favorisce la
digestione.
, da non scordare il cibo sminuzzato che favorisce la
digestione.
Una volta raggiunta la maturitŕ sessuale, l'attivitŕ
dell'ovaio diventa ciclica. Nella gonade di una gallina sono costantemente
presenti 4-6 oociti nella terza fase di accrescimento tra loro
legati in via gerarchica. Di norma ogni giorno un follicolo compie il proprio
ciclo ovulatorio e un altro follicolo viene richiamato dalla schiera di oociti
in attesa di intraprendere l’accrescimento. Cosě l’ovulazione č
quotidiana e ogni ciclo possiede una pausa. La durata della sequenza č
caratteristica della razza e talvolta dell'individuo.
Č ovvio che le sequenze piů lunghe sono particolarmente ricercate nelle
razze allevate per la produzione di uova. Le analisi ormonali e le indagini
sperimentali hanno permesso di chiarire, almeno in linea generale, i
meccanismi endocrini che sovrintendono all’attivitŕ ciclica dell'ovaio.
La nuova ovulazione segue di 30
minuti
la deposizione appena avvenuta, salvo che l’uovo sia stato deposto nel tardo
pomeriggio. In tal caso le nuova ovulazione viene rinviata al primo mattino
del giorno successivo. Da notare tuttavia che l’ovulazione non dipende dal
fatto meccanico dello sgravarsi, poiché si verifica a intervalli regolari
anche se l’uovo precedente č stato tolto prematuramente dall’utero.
L’ora della deposizione dipende dall’illuminazione:
una gallina che fa l’uovo ogni 26
ore,
deporrŕ alle 8, 10, 12, 14, 16, e deporrŕ per 5
giorni
di seguito, quindi riprenderŕ a deporre la mattina di dopodomani. Se si
rovesciano le condizioni d’illuminazione -
oscurando il giorno e illuminando la notte - si ottiene uno spostamento
di mezza giornata sull’ora della deposizione. Se l’illuminazione č
continua la gallina puň ovulare e deporre in qualunque ora del giorno e della
notte. Dal punto di vista ormonale l’ovulazione č provocata dall’ormone
luteinizzante ed č arrestata dalla prolattina. Se la deposizione č
troppo precoce rispetto alla norma, si osservano uova con guscio molto
sottile, addirittura solo membranaceo.
Da notare inoltre che la gallina č in grado di deporre
uova feconde per un certo tempo dopo il suo isolamento:
da 15-20
giorni
sino a 32 giorni dall’ultima copula.
1.3.c. Regolazione ormonale dell’ovulazione
Lo sviluppo dei follicoli
ovarici č indotto dalle gonadotropine ipofisarie: FSH
(follicolostimolante) e LH (luteinizzante). Tuttavia il
ruolo specifico di questi due ormoni č ancora poco chiaro, ma si ritiene che
l’LH induca l’attivitŕ steroidogenica della gonade e sia primariamente
responsabile dell'ovulazione, mentre pare che l’FSH causi soprattutto l’accrescimento
dei follicoli. L’immissione in circolo della quantitŕ di gonadotropine
necessarie a indurre la maturazione dell'ovaio nell'animale in accrescimento
č determinata da specifici fattori rilascianti elaborati dai nuclei
ipotalamici e inviati all'adenoipofisi attraverso il sistema vascolare
ipotalamo-ipofisario. Questa connessione neuroendocrina acquista particolare
significato se si tiene conto che l’ipotalamo riceve afferenze da altri
centri nervosi cui fanno capo diversi sistemi sensoriali capaci di percepire
parecchie modificazioni chimico-fisiche dell'ambiente, come luce e
temperatura.
L’FSH, con il contributo dell'LH, avvia l’accrescimento
del follicolo nell'animale che si approssima alla prima ovulazione. Sotto l’influsso
dell'LH il follicolo elabora estrogeni
che, immessi in circolo, raggiungono il fegato e vi inducono la sintesi dei
composti destinati a formare il vitello. Man mano che il follicolo si
accresce, esso produce progesterone che attraverso l'ipotalamo
č in grado di stimolare, un giorno prima della prevista ovulazione, un centro
ciclico dal quale dipende il rilascio episodico di una notevole quantitŕ di
LH (effetto feedback
[3]
positivo). Il follicolo intanto č diventato particolarmente sensibile all'LH
e come risposta a questo ormone acquista la capacitŕ di ovulare. In seguito
all'ovulazione la sintesi di progesterone decresce rapidamente e quindi il
ciclo si ripete, interessando il secondo follicolo della gerarchia e cosě
via.
Da quanto detto risulta evidente che a livello ipotalamico
vanno distinti due centri funzionali. Il primo, spesso indicato come centro tonico, sostiene il rilascio di
gonadotropine necessarie allo sviluppo dei follicoli, mentre l’altro, il centro
ciclico, interviene solo in rapporto all'ovulazione ed č
stimolato dal progesterone.
Le determinazioni dei livelli ematici di LH e di
progesterone sembrano rendere plausibile il modello ormonale illustrato, ma,
prima dell'aumento della gonadotropina, si osserva anche un significativo
incremento del testosterone circolante. Va aggiunto che la sensibilitŕ
ipotalamica al progesterone deve essere opportunamente risvegliata dagli
estrogeni perché si abbia un adeguato rilascio di LH. Recentemente č stato
suggerito che vari corticoidi potrebbero in qualche modo interferire nella
sequenza.
1.3.d. Influenze esogene
Molti fattori possono
interferire in modo abbastanza vario con lo sviluppo e con l’attivitŕ della
gonade. A titolo d'esempio: l’insufficiente alimentazione
determina un ritardo dello sviluppo postnatale che si riflette inevitabilmente
sulla maturazione sessuale.
Piů diretti e critici sono gli effetti del fotoperiodo,
cioč delle ore di luce giornaliere. Negli uccelli che vivono in regioni
temperate l’incremento del fotoperiodo in primavera e il decremento nella
tarda estate o nell'autunno agiscono da regolatori per l’attivitŕ delle
gonadi. In linea generale l’allungamento del fotoperiodo si comporta da
stimolante, mentre il suo accorciamento funziona da inibente. Tali fenomeni
sono abbastanza evidenti negli animali allo stato libero, mentre acquistano
minor valore nelle specie domestiche, in particolare nel pollo, che č stato
selezionato da soggetti viventi ai tropici. Ciň nonostante, in tutti gli
uccelli domestici l’incremento del fotoperiodo puň anticipare la comparsa
della maturitŕ sessuale, mentre la riduzione č in grado di ritardarla. Inoltre
i
regimi fotoperiodici interferiscono con la carriera riproduttiva. Ad
esempio, fotoperiodi brevi e costanti di 6 ore diminuiscono la produzione totale di uova; nel
tacchino e nella quaglia sono richieste almeno 10 ore
giornaliere di luce per avere uno sviluppo normale della gonade e una buona
produzione di uova.
Nella pratica zootecnica sono stati introdotti particolari
programmi di luce da somministrare in vari periodi della vita, al fine di
contemperare le esigenze di un buon sviluppo corporeo con una sufficiente resa
produttiva. Esiste ormai la possibilitŕ di scegliere tra un numero di uova
elevato e basso, in questo caso fornite di guscio piů robusto.
Il meccanismo d’azione della luce implica l’interessamento
sia di centri nervosi che dell'epifisi, i quali vanno a regolare i centri
ipotalamici produttori dei fattori rilascianti.
2. Ovidutto sinistro
Nella maggioranza degli uccelli si sviluppa solo l’ovidutto
sinistro. Il destro si atrofizza precocemente e nell'adulto se ne possono
osservare dei residui. L'ovidutto presenta variazioni di volume imputabili
alla riproduzione, molto evidenti nei riproduttori stagionali, mentre nella
gallina questo aspetto riveste scarsa importanza. In fase ovulatoria l’ovidutto
svolto della gallina ha una lunghezza di 50-80 cm
con un peso di oltre 70 grammi.
L'ovidutto č un condotto convoluto a pareti spesse posto
tra l’ovaio e la cloaca, in rapporto superiormente con l’ovaio e,
caudalmente a questo, con il rene sinistro e con la volta della cavitŕ
pelvica; stretti rapporti si stabiliscono anche con il sacco aereo addominale
sinistro.
In direzione anteroposteriore presenta cinque tratti,
distinguibili per i caratteri morfologici e per la struttura istologica:
infundibolo, magnum, istmo, utero e vagina. Tutto l’ovidutto č sospeso alla
volta della cavitŕ addominale da un legamento peritoneale che lo ingloba e
che si continua ventralmente.
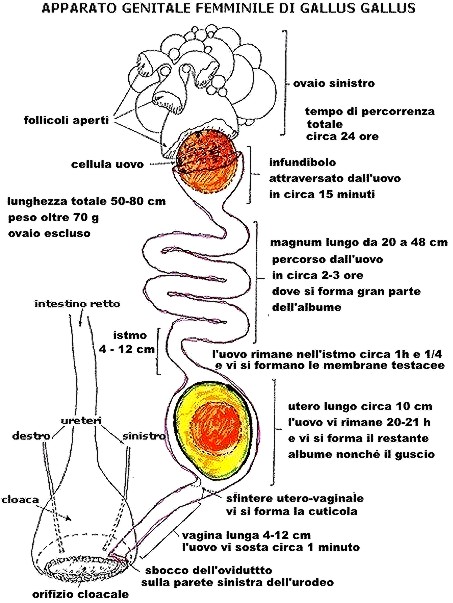
Fig. VIII. 4 - Schema di ovidutto di gallina
Non sono rispettate le reali lunghezze dei vari segmenti
Infundibolo
- Rappresenta l’estremitŕ craniale dell'ovidutto. Č paragonabile a un
largo imbuto schiacciato lateralmente e disposto in prossimitŕ del polo
posteriore dell'ovaio. Possiede dei prolungamenti nastriformi, le fimbrie, che
si spingono tra i follicoli in accrescimento per rendere piů agevole la
raccolta degli oociti al momento dell'ovulazione. Nel punto di giunzione con
il segmento successivo si trovano delle strutture tubulari, le fossette
ghiandolari, dove vengono immagazzinati gli spermatozoi.
Magnum
- Come dice il nome č il tratto piů lungo dell'ovidutto: nella gallina varia
da 20
a
48
cm.
Ha pareti spesse e descrive diverse anse. Lo spessore della parete č dovuto
al gran numero di ghiandole che diminuiscono nell'ultimo tratto, in
prossimitŕ dell'istmo.
Istmo
- Appare come una breve e ristretta regione lunga 4-12 cm
nella gallina; si distingue per il colore piů chiaro.
Utero
- Appare come un’espansione a forma di sacco e nella gallina č lungo circa 10
cm.
Le pliche della mucosa hanno andamento longitudinale, trasversale e obliquo,
sě da conferire alla superficie interna un aspetto irregolare per la
sporgenza di lamelle alte alcuni millimetri. Anche nel punto di giunzione tra
utero e vagina si trovano delle formazioni tubulari che servono a
immagazzinare gli spermatozoi, le fossette spermatiche.
Vagina
- Rappresenta il tratto terminale dell'ovidutto, diretta continuazione dell’utero:
caudalmente si apre sulla parete sinistra dell'urodeo che č un tratto della
cloaca. Lunga 4-12 cm nella gallina, assume una
caratteristica forma a S.
2.1. Struttura dell’ovidutto
Nella parete dell'ovidutto si
distinguono diversi strati, due dei quali sono muscolari, ma a noi interessano
essenzialmente quelli deputati all’attivitŕ secretoria. L’epitelio
mucoso, costituito da cellule cilindriche, presenta sia elementi ciliati che
secernenti. La proporzione relativa dei due tipi cellulari č
variabile, ma in genere 6 cellule secernenti circondano una cellula ciliata.
La corrente prodotta dalle cilia č diretta verso la vagina, ma nel magnum si
trovano regioni in cui il movimento č inverso e le cellule ciliate
favorirebbero la progressione degli spermatozoi. Le cellule secernenti, di
tipo caliciforme, elaborano soprattutto mucine che vengono liberate durante il
passaggio dell’uovo.
Alla mucosa sono associate
ghiandole pluricellulari che hanno forma di docce ghiandolari o di vere
ghiandole tubulari. Le prime, disposte essenzialmente nell'infundibolo,
appaiono come solchi di elementi secernenti compresi tra le pliche mucose
secondarie. Le ghiandole tubulari si aprono nelle depressioni delimitate dalle
pliche secondarie e sono diffuse in tutto l’ovidutto. Le loro cellule
secernenti sono alte e ricche in granuli di secreto liberati con meccanismo di
secrezione apocrina
[4]
.
Nella vagina le ghiandole sono limitate al tratto iniziale
e acquistano particolare importanza in quanto sarebbero in grado di garantire
la vitalitŕ degli spermatozoi per oltre 14
giorni;
il loro secreto č ricco in colesterolo e suoi esteri.
2.2. Fisiologia dell’ovidutto
La funzione dell'ovidutto
consiste essenzialmente nel fornire alla cellula uovo un insieme di involucri
concentrici: l’albume, le membrane testacee e il guscio, oltre a consentire
la progressione dell'uovo mediante opportune contrazioni peristaltiche. Del
guscio abbiamo giŕ parlato estesamente nel capitolo riguardante le uova blu
dell’Araucana (vol.I – IX.4.)
2.3. Composizione dell’albume
Nell’uovo deposto l’albume -
o bianco d’uovo - si presenta come una sostanza vischiosa traslucida il cui
peso corrisponde a circa 2/3
dell’intero
uovo, formata quasi esclusivamente da proteine e acqua in proporzione 1:8.
Vi si trovano disciolti anche zuccheri e minerali. Nell’uovo ormai deposto l’albume
non č omogeneo, essendo costituito da strati distinti che procedendo dal
tuorlo sono i seguenti:
·
calazifero
- 2,7%
-
dal quale partono 2 cordoncini ritorti, le calaze
[5]
,
orientate verso ciascun polo dell’uovo e che hanno la funzione di mantenere
l’ovulo in posizione centrale
·
sottile
interno - 17,3% - fluido
·
gelatinoso
- 57% - denso per la presenza di
mucina
·
sottile esterno
- 23% - di nuovo fluido.
Il rivestimento albuminoso viene applicato al tuorlo
durante il passaggio attraverso l’ovidutto. I meccanismi che ne controllano
la sintesi sono noti solo in parte.
Come abbiamo giŕ accennato,
disciolti nella componente acquosa dell'albume si trovano anche zuccheri e
ioni minerali; molti di questi sono legati a proteine.
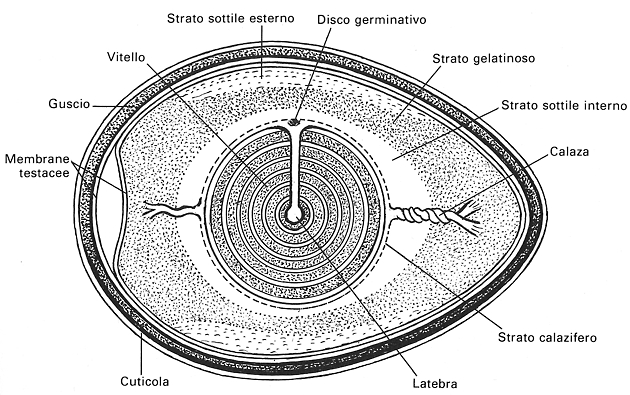
Fig. VIII. 5 - L’uovo: disegno schematico che mette bene in evidenza la struttura dell’albume.
|
Proteine
dell'albume dell’uovo di gallina da Botte & Pelagalli |
|||||
|
Proteine |
% |
g/uovo |
Peso |
% |
Proprietŕ |
|
Ovalbumina |
54 |
2,05 |
46.000 |
3 |
|
|
Ovotransferrina |
13 |
0,49 |
76.000÷86.000 |
2 |
lega
Fe, Cu,
Mn, Zn; batteriostatica |
|
Ovomucoide |
11 |
0,42 |
28.000 |
22 |
inibisce
la tripsina |
|
Globulina
G2 |
4 |
0,03 |
fra 36.000 |
|
|
|
G3 |
4 |
0,16 |
e 45.000 |
|
|
|
Lisozima |
3,5 |
0,13 |
14.300÷17.000 |
2 |
lisa
i batteri |
|
Ovomucina |
1,5÷2,9 |
|
|
19 |
emoagglutinazione
antivirale |
|
Flavoapoproteina |
0,8 |
0,03 |
32.000÷36.000 |
14 |
lega
la riboflavina |
|
Ovomacroglobulina |
0,5 |
0,02 |
760.000÷900.000 |
9 |
|
|
Ovoglicoproteina |
0,5÷1 |
|
24.400 |
16 |
|
|
Ovoinibitore |
0,1÷1,5 |
|
46.000 - 49.000 |
6 |
inibisce
la proteasi |
|
Avidina |
0,05 |
0,02 |
68.300 |
8 |
lega
la biotina |
|
non
identificate |
circa 0,8 |
|
|
|
|
La maggior parte delle proteine
č secreta dalle ghiandole del magnum, attivitŕ che č controllata dagli
ormoni sessuali. Gli estrogeni e il testosterone stimolano l’accrescimento
dell'ovidutto e la sua attivitŕ secernente, che tuttavia necessita del
progesterone per raggiungere il potenziale normale. Almeno una proteina, l’avidina,
č sintetizzata solo dopo stimolazione con progesterone. Altre componenti,
come l’ovotransferrina, gli ioni e l’acqua, derivano direttamente dal
sangue e passano nel secreto con meccanismi poco noti. L'attivitŕ sintetica
delle ghiandole dell'ovidutto č enorme, se si pensa che il corredo di albume
di un uovo viene prodotto nel giro di 24 ore.
Secondo alcuni questa funzione viene stimolata anche dal tuorlo stesso man
mano che percorre l’organo.
3. Ovodeposizione
I tempi di percorrenza
dell'ovidutto sono caratteristici per ciascuna regione, anche in rapporto alle
funzioni esplicate da ciascuna. Nel pollo l’infundibolo č percorso in 15
minuti,
il magnum in 2-3 ore, l’istmo in un'ora e un quarto, l’utero in 20-21
ore
e la vagina in circa un minuto. Il tempo totale č di circa 24
ore.
I valori cambiano leggermente in altre specie: cosě il tempo totale č di 26
ore nel tacchino e di 25 ore nella quaglia.
I meccanismi di avanzamento dell'uovo nell'ovidutto sono
poco noti. Le differenze nella velocitŕ di percorrenza dei vari tratti
richiedono una fine regolazione che č stata di volta in volta imputata al
sistema nervoso o all'apparato endocrino.
Poiché l’attivitŕ dell'infundibolo aumenta ancor prima
che si verifichi l'ovulazione, si č supposto l’intervento degli ormoni
ovarici, ma si č ancora lontani da una dimostrazione conclusiva. La
sensibilitŕ dell'ovidutto all'acetilcolina e all'adrenalina, particolarmente
notevole per l'utero, ha fatto ipotizzare dei sistemi di controllo nervosi.
In effetti, ricerche citologiche dimostrano che l’innervazione dell'ovidutto
č molto complessa e i diversi tratti presentano qualche particolaritŕ
distintiva.
Parimenti complicato appare il sistema che controlla la
deposizione. Il follicolo postovulatorio interverrebbe con un ormone, peraltro
da identificare, il quale allunga i tempi di percorrenza dell'ovidutto e
ritarda la deposizione. Anche il sistema nervoso, comunque, č implicato in
questa funzione: la stimolazione del telencefalo ritarda la deposizione,
quella dell'ipotalamo l’anticipa. Va precisato, perň, che le sostanze
attive sulle funzioni del sistema nervoso vegetativo danno risultati poco
congruenti con questa ipotesi.
In merito alla regolazione ormonale, per un certo tempo si
č ritenuto che il progesterone fosse l’agente stimolante delle rapide
contrazioni uterine, cui consegue la deposizione. Successivamente s’č visto
che diversi ormoni possono accentuare le contrazioni dell'utero, compresi gli
estrogeni, e che il livello ematico di progesterone raggiunge i valori piů
alti ben 12 ore prima dell'espulsione dell'uovo.
Un ruolo determinante nel fenomeno espulsivo potrebbe
essere svolto dagli ormoni della neuroipofisi, ossitocina
[6]
e arginina-vasotocina,
i quali stimolano i muscoli lisci. Il loro intervento č reso ancora piů
plausibile dal fatto che essi diminuiscono nell’ipofisi e aumentano nel
sangue appena prima della deposizione. Tuttavia la distruzione enzimatica
dell'ossitocina circolante non sembra interferire con la deposizione.
Anche le prostaglandine
[7]
sono intimamente coinvolte nel processo di ovodeposizione e la loro
concentrazione ematica aumenta in modo significativo al momento della
deposizione per subire una drastica caduta nel periodo immediatamente
successivo. Inoltre l’ovodeposizione puň venir differita attraverso un’immunizzazione
passiva nei confronti della prostaglandina PGE1,
mentre puň venir anticipata dall’iniezione di prostaglandina. Le
prostaglandine esogene sono molto piů potenti dell’ossitocina nell’indurre
l’ovodeposizione, mentre il blocco farmacologico della sintesi
prostaglandinica inibisce contemporaneamente l’uscita dell’uovo all’esterno.
La prostaglandina PGF2a
aumenta le contrazioni muscolari e la pressione in tutti i segmenti dell’ovidutto,
mentre la PGE1 e la PGE2
aumentano le contrazioni in tutti i segmenti eccetto che a livello della
vagina dove inducono il rilasciamento della muscolatura. Queste risposte
divergenti rappresentano un ottimo adattamento fisiologico alle due classi di
prostaglandine, in quanto al momento della progressione dell’uovo nei tratti
finali dell’ovidutto si verifica la possibilitŕ del suo passaggio in
cloaca. Pertanto secondo Hertelendy (1984) l’azione combinata di PGF2a,
PGE1
e PGE2
potrebbe rappresentare il meccanismo regolatore finale dell’ovodeposizione.
Dal momento che un discreto numero di sostanze sembra
interferire con il tempo di deposizione, si č portati a credere che questo
processo venga di norma regolato da numerosi fattori tra loro in cooperazione.
L'intervento della vagina sembra essere di minore importanza, anche se la sua
dilatazione meccanica puň essere all'origine dei riflessi che portano al
rilassamento della muscolatura a livello della giunzione uterovaginale e
quindi, successivamente, alla contrazione peristaltica dell'utero.
4. Deposizione di due uova al giorno
La deposizione da parte di una
gallina di due uova al giorno č
un argomento al quale abbiamo giŕ accennato nel capitolo storico, ma ora lo
riprendiamo sotto un profilo prettamente scientifico, facendo un rapido
excursus della letteratura in merito.
Drew (1907) e Curtis (1914)
descrissero la deposizione di due uova giornaliere da parte dello stesso
soggetto, senza peraltro accennare ad anomalie del guscio. Altri studiosi che
piů tardi descrissero la doppia deposizione giornaliera rimasero
particolarmente colpiti dal fatto che il secondo uovo aveva una forma anomala
(Scott, 1940; Grau & Kamei, 1949;
Weiss & Sturkie, 1952; Foster, 1970).
L’anomalia del primo uovo deposto non č cosě evidente come lo č per il
secondo. Grau & Kamei avevano notato che il primo uovo possedeva un guscio
spesso e ruvido come la sabbia, e un guscio piů spesso era stato rilevato
anche da Foster. Uova la cui superficie si presentava rugosa come la carta
vetrata furono riferiti anche da Jaap & Muir (1968),
ma essi non riuscirono a stabilire se questa caratteristica riguardasse il
primo oppure il secondo uovo. Scott notň che un uovo di consistenza
membranacea con un’area appiattita viene causato dal contatto con un altro
uovo a livello dell’ovidutto. L’ipotesi sostenuta da Middelkoop (1971)
circa l’origine delle peculiaritŕ delle due uova č la seguente: il primo
uovo rimane nell’utero al di lŕ dello scadere del tempo per la sua
deposizione e il successivo uovo, scendendo lungo l’ovidutto, giunge in
utero e preme contro l’uovo quivi ritenuto. Essendo il secondo uovo in uno
stato di tensione ancora insufficiente, la pressione contro l’altro uovo ne
causa la caratteristica malformazione.
Le madri dei broiler non solo depongono meno uova delle
galline selezionate per ovodeposizione, ma le loro uova presentano una maggior
percentuale di anomalie. Jaap & Muir hanno notato che durante i primi sei
mesi di deposizione le galline da broiler producevano uova con doppio tuorlo
con frequenza doppia rispetto ad altre popolazioni di galline, e che esse
possono deporre due uova al giorno con anomalie del guscio, alcune delle quali
si presentano membranose.
Middelkoop (1971) intraprese
uno studio sistematico di 624 femmine di una linea pura di
Plymouth Rock bianca, tenute in gabbie separate. A un certo punto dieci
galline tra quelle che avevano deposto regolarmente due uova nell’arco di 24
ore
furono trasferite in batterie al fine di permettere la registrazione
automatica del momento della deposizione.
Cosě a Middelkoop č stato possibile procedere alla
determinazione delle caratteristiche delle singole uova. Il primo di un paio
di uova possiede un guscio duro, cui si aggiunge un’ulteriore calcificazione
rugosa, che talora interessa l’intera superficie, ma abitualmente si
presenta sotto forma di banda, che ha un decorso piů o meno longitudinale. La
vera cuticola - che č lo strato organico piů esterno dell’uovo - č
presente, ma si trova al di sotto dello strato rugoso. La presenza della
cuticola vera al di sotto della deposizione calcarea aggiuntiva sta a
significare che questo primo uovo č completamente formato e che la
calcificazione si č arrestata quel tanto da permettere l’apposizione della
cuticola. Dal momento che nelle uova a guscio marrone lo strato calcareo
aggiuntivo si presenta bianco, ciň implica pure che la formazione dell’involucro
si č arrestata per poi riprendere.
Il secondo uovo spesso si presenta con un guscio piů o
meno molle e per lo piů č caratterizzato da un’area appiattita che č piů
sottile del rimanente guscio ed č circondata da un anello di guscio
raggrinzito, piů vicina al polo acuto che a quello ottuso. Questo fenomeno
puň essere definito come guscio compresso da un lato, e non uovo
troncato come proposto da Romanoff & Romanoff (1949), in
quanto la prima definizione riassume sia la forma che la causa che ha condotto
all’alterata morfologia.
Si č potuto osservare che quelle galline che spesso
depongono due uova al giorno mostrano questa caratteristica in modo del tutto
regolare. Impiegando un registratore automatico, Middelkoop ha potuto rilevare
che il momento della deposizione del primo uovo corrisponde abitualmente all’inizio
della notte fino a dopo mezzanotte. In media il secondo uovo viene deposto 15 ore piů tardi, ma con un ampio margine di
variabilitŕ, in quanto la deviazione standard assomma a circa 5
ore
e mezzo.
Un dato certo al quale Middelkoop č giunto attraverso radiografie č che il cosiddetto primo uovo permane in utero molto piů
a lungo del solito. Un altro dato interessante, perň di tipo comportamentale
a conferma di quello radiologico, consiste nel fatto che si possono osservare
galline che vanno nel nido-trappola al giusto momento della deposizione, ma
senza risultato; durante la sera o durante la successiva notte un primo uovo
viene deposto al suolo, per cui ciň sta a significare che il primo uovo č
rimasto nell’ovidutto molto piů a lungo del normale. Dopo la deposizione
del primo, il secondo uovo cerca di adeguare la sua area compressa a una forma
abituale mentre si trova in utero, dando origine talora, anziché a un’area
appiattita, a un rigonfiamento del contorno, disposto per lo piů in
corrispondenza della zona equatoriale.
La causa piů verosimile della doppia deposizione
giornaliera consisterebbe in un troppo breve intervallo di tempo
fra due successive ovulazioni. Sappiamo che in una sequenza
normale l’ovulazione segue di mezz’ora la deposizione, ma si tratta di un
intervallo che non sempre č costante, in quanto puň oscillare fra 30
minuti
e un’ora e mezza. In linea di massima lo stimolo all’ovulazione - che
determina l’intervallo di tempo fra due ovulazioni successive - proviene dal
coordinamento tra il differente grado di maturitŕ del follicolo rispetto agli
altri follicoli e il ritmo nictemerale. Middelkoop ha potuto concludere che l’ovulazione,
in condizioni di illuminazione naturale, puň verificarsi in un momento che va
da 10
ore
a partire dall’imbrunire fino a 10 ore dopo l’alba. In pratica, se
prendiamo ad esempio il mese di marzo e il 45° parallelo nord, l’ovulazione
puň verificarsi dalle 4 del mattino alle 4 del
pomeriggio.
Due ovulazioni contemporanee o in rapida successione danno
luogo a un uovo con doppio tuorlo. Ma, se un’ovulazione successiva si
verifica prima che il precedente uovo venga deposto, ecco che il primo uovo,
per ragioni puramente fisiologiche connesse al funzionamento dell’ovidutto,
rimane piů a lungo in utero, rivestendosi di ulteriore guscio. In questo caso
le due ovulazioni sono separate, in media, da un intervallo di sole 20-22
ore.
5. Struttura dell’uovo
Abbiamo parlato della natura
chimica dei componenti dell’uovo e dei meccanismi preposti alla loro
sintesi. Vediamo ora come l’uovo č organizzato internamente.
Nell'uovo deposto bisogna
distinguere:
·
la
cellula uovo, o tuorlo, o rosso d'uovo
·
l’albume,
o bianco d'uovo
·
il
guscio, che comprende anche le membrane testacee.
Anche se l’uovo acquista la
forma e il volume caratteristici della specie, la proporzione relativa dei
costituenti č abbastanza costante.
|
Percentuale
dei componenti
da Botte & Pelagalli |
|||
|
Componenti |
Guscio |
Albume |
Cellula uovo |
|
acqua |
1 |
88,5 |
47,5 |
|
proteine |
4 |
10,5 |
17,4 |
|
lipidi |
|
|
33 |
|
carboidrati |
|
0,5 |
0,2 |
|
ioni
minerali |
95 |
0,5 |
1,1 |
|
altri |
|
|
0,8 |
5.1. Cellula uovo
La cellula uovo ha forma
subsferica e in un uovo medio di 58 g
pesa circa 19 g con un diametro di 35-40
mm
e un volume di 16-17 cmł. Consta di tre parti: la membrana vitellina,
il disco germinativo e il vitello.
La
membrana vitellina avvolge il tuorlo, ma va distinta dalla struttura omonima
della cellula uovo contenuta nel follicolo ovarico. Di questa, infatti,
restano solo dei frammenti in quanto si dissolve in parte prima
dell'ovulazione. La membrana vitellina presenta una struttura complessa,
essendo formata da due strati di derivazione ovarica - i resti della membrana
vitellina e la membrana perivitellina - e da due strati probabilmente prodotti
dall'infundibolo e quindi derivati dall'albume: la membrana continua e la
membrana extravitellina.
Nell'ambito della membrana perivitellina
si osservano delle strutture fibrose ampiamente anastomizzate a rete e
orientate parallelamente alla superficie dell’uovo spesse 2 mm.·La membrana
continua, spessa 50-100 mm,
ha un aspetto granulare, mentre al suo esterno la membrana
extravitellina appare come una rete di sottili fibrille spessa 3-9
mm.
Nel suo complesso la membrana vitellina ha una notevole
resistenza, specialmente verso la regione appuntita dell'uovo. Č costituita
per l'80% da acqua; il 90% dei
solidi č rappresentato da proteine e il 3% da
lipidi. In linea di massima le sue parti piů interne somigliano al collagene,
mentre quelle piů esterne sono abbastanza simili all'albume.
La permeabilitŕ della membrana č alquanto
caratteristica, in quanto si lascia attraversare quasi esclusivamente da
acqua, che viene scambiata fra tuorlo e albume.
Il disco germinativo č una piccola parte
della cellula uovo che a causa della sua minore densitŕ occupa il polo
animale. Nell'uovo fecondato consta di una massa di 40.000-60.000 cellule
derivate dalla divisione ripetuta dello zigote e prende il nome di discoblastula
o blastoderma, che appare come un dischetto di colore grigio chiaro del
diametro di 4 mm che riposa sulla componente bianca del vitello. Nel
caso l’uovo non sia stato fecondato, il disco germinativo č costituito da
citoplasma e dal nucleo femminile in degenerazione e il suo diametro č di
circa 3,5 mm.
Il vitello rappresenta la componente
maggiore della cellula uovo ed č strutturato in due porzioni facilmente
distinguibili. Al di sotto del disco germinativo si trova una struttura conica
di aspetto chiaro che si approfonda verso il centro della cellula, ove termina
con una porzione sferica di circa 5 mm
di diametro, la latčbra
[8]
,
composta di vitello bianco, e che rappresenta appena l'1-2%
del
totale. Attorno alla latebra si dispone il vitello giallo.
Si č molto discusso se questo sia presente in strati sovrapposti, dal momento
che in molti casi appare come un'alternanza di fasce giallo-rosse di diversa
intensitŕ. Oggi pare assodato che tale aspetto non rispecchi una particolare
disposizione spaziale, ma dipenda dalla ricchezza in xantofilla della dieta.
5.2. Albume
Anche l’albume presenta una
tipica organizzazione macroscopica in quanto non č fisicamente omogeneo. Si
distinguono due calaze e diversi strati.
Le calaze si dipartono da ciascun polo
della cellula uovo e sono dirette secondo l’asse maggiore del guscio. Si
tratta di strutture cordoniformi avvolte su se stesse. Verso il polo ottuso si
dirige una sola calaza, mentre dall'altro lato ne esistono due tra loro
intimamente ritorte. Originano a livello dello strato calazifero e terminano
da ciascun lato nella regione dei legamenti dell'albume.
Lo strato calazifero č cospicuo, ha
struttura omogenea ed č a stretto contatto con la membrana vitellina della
cellula uovo.
Gli strati sottili interno ed esterno appaiono
piuttosto fluidi e omogenei; nella loro compagine non si distinguono fibre,
anche se sono presenti mucine.
Lo strato gelatinoso č molto ricco in
ovomucina che conferirebbe l’aspetto di gel denso. A ciascuna estremitŕ
dell'asse maggiore dell'uovo si prolunga nei legamenti dell'albume connessi
mediante mucine alle membrane testacee. Si č molto discusso
sull'organizzazione submicroscopica di questa porzione dell'albume e
soprattutto č dibattuta la presenza o meno di fibre.
5.3. Membrane testacee
Si tratta di due membrane
leggermente elastiche e biancastre accollate fra loro, salvo che in
corrispondenza del polo ottuso dell'uovo dove formano la camera d'aria.
Quest'ultima si genera per il rapido raffreddamento dell’uovo dopo la
deposizione, che porta alla penetrazione di aria e allo scollamento delle due
lamine. Ciascuna membrana č composta da piů strati.
La membrana interna č organizzata in almeno
tre piani di fibre orientate in modo differente, per uno spessore totale di
circa 22 mm.
La membrana esterna č piů complessa. Ha
uno spessore di circa 50 mm
e possiede fibre brevi e spesse, poste in almeno 6 piani
e orientate ad angolo retto tra piano e piano.
Per la struttura, la
composizione chimica e la pigmentazione del guscio, si rimanda alla parte
storica riguardante l’Araucana (vol.I – IX)
6. Uovo di gallo - Uovo senza giallo - Uovo bianco
Il tuorlo č in grado di
stimolare meccanicamente la produzione dell’albume, che infatti puň essere
ottenuto anche con sfere d’ambra, d’osso e di cera deposte all’inizio
dell’ovidutto. Anche le feci deviate artificialmente verso l’utero vengono
ricoperte d’albume. Le uova senza tuorlo d’origine spontanea dimostrano
tuttavia che un corpo estraneo non č di capitale importanza per la loro
formazione; tali uova sono costituite solo da albume e guscio, sono anche
dette uova di gallo, e sono frequenti
verso
la fine del periodo depositivo.
Si pensava che le uova potessero venire deposte anche dal
gallo, che da tali uova talora potesse nascere un basilisco. Di queste
fantasie parleremo quando analizzeremo l’uovo secondo Aldrovandi.
Le uova senza tuorlo, sfrondate delle
fantasie del passato - come puntualizzeremo a proposito dell’uovo secondo
Aldrovandi - vengono fondamentalmente distinte in due classi:
·
uova
contenenti parassiti o altri corpi estranei con funzioni di stimolo meccanico
·
uova
prive di qualsiasi incluso cui possa essere attribuita la produzione abnorme
di albume.
In questo secondo caso l’ipotesi piů accreditata č
quella secondo cui il solo accumulo di albume nella parte albuminifera del
condotto č in grado di stimolare la formazione del rivestimento calcareo.
Esiste tutta una serie di uova anomale senza alcun
addentellato con miti e fantasie. Infatti esistono uova
giganti per ipertrofia dell’albume o per coesistenza di 2
o
piů tuorli; esistono uova nane avitelline ![]() ,
uova a guscio sottile, a guscio spesso, con 2-3 o
piů gusci, uova contenenti parassiti, ghiaia, piume. Esistono poi le vere
uova doppie - ovum in ovo o uovo matreshka
,
uova a guscio sottile, a guscio spesso, con 2-3 o
piů gusci, uova contenenti parassiti, ghiaia, piume. Esistono poi le vere
uova doppie - ovum in ovo o uovo matreshka![]() - formate da un uovo che ne
racchiude un altro.
- formate da un uovo che ne
racchiude un altro.
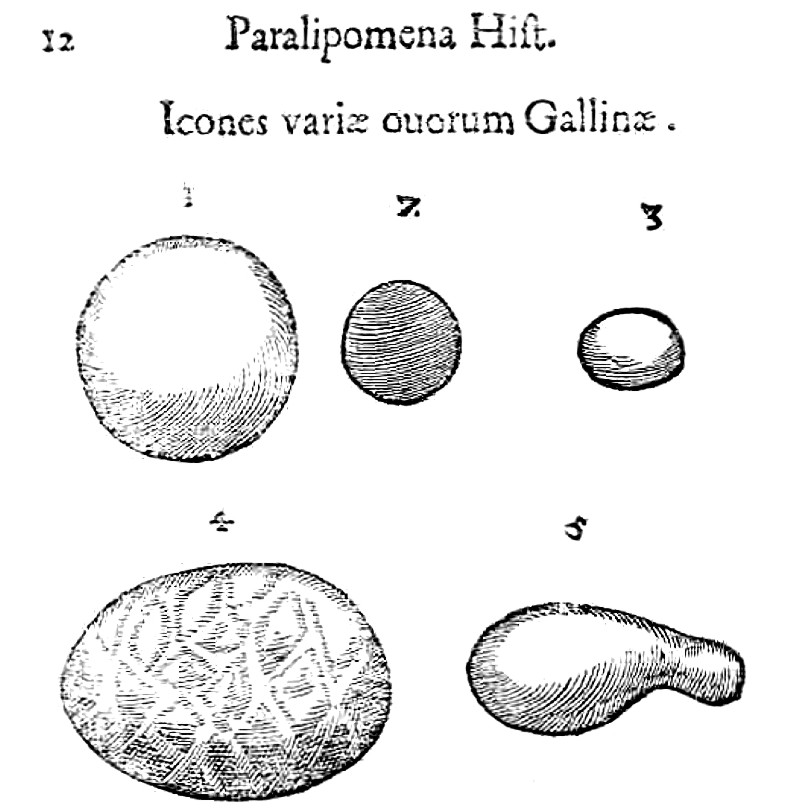
Insolite
uova di gallina
dai Paralipomena
accuratissima historiae omnium animalium (1642) di Ulisse Aldrovandi
L'annessa didascalia riporta quanto segue:
1 – Ovum gallinae rotundum instar pilae
Uovo di gallina rotondo come una palla
2 – Ovum aliud gallinae parcum rotundum cortice
durissimo
Altro uovo di gallina, piccolo, rotondo, dal guscio estremamente duro
3 – Ovum gallinae magnitudine ovi columbini
Uovo di gallina grande come un uovo di piccione
4
– Ovum gallinae monstrificum, nempe undosum, et
rugosum
Uovo di gallina mostruoso, ossia a superficie ondulata e rugosa
5 – Ovum gallinae monstrosum nempe figura
cucurbitae perticalis
Uovo di gallina mostruoso, cioč foggiato come una zucca che si arrampica
sulle pertiche
7. Ovum in ovo
Uovo matrioska 


Matrioska della Bielorussia - 1996

La produzione di uova doppie č
piuttosto rara e di esse si occupa quella branca scientifica denominata teratologia,
che in greco significa mostruositŕ. Forse la piů antica descrizione
di uova doppie risale al 1250 a opera di Alberto Magno![]() che
in
De animalibus I,81 fa
menzione di un uovo con due gusci
che
in
De animalibus I,81 fa
menzione di un uovo con due gusci![]() :
:
Ego tamen iam vidi ovum gallinae, quod habuit duas testas, unam intra aliam, et in medio duarum testarum habuit albuginem, et intra interiorem etiam non fuit nisi albugo, et fuit ovum parvum, totum rotundum ad modum sperae. Sed hoc erat unum de naturae peccatis et monstris.
Tuttavia giŕ vidi un uovo di gallina che aveva due gusci, uno dentro all'altro, e tra i due gusci c'era dell'albume, e all'interno di quello che stava dentro altro non c'era che albume, ed era un uovo piccolo, completamente rotondo come una sfera. Ma questo apparteneva a uno degli errori e delle anomalie della natura.
Dai casi descritti col volgere dei secoli
risulta che vi possono essere diversi tipi di uova doppie, sia per forma che
per grandezza. L’uovo esterno puň essere piů piccolo o piů grande del
normale. Quello interno č invece generalmente piů piccolo del normale. L’uno
o l’altro puň essere privo di guscio, o averlo appena accennato, oppure
sottilissimo. Spesso le uova doppie presentano forma anomala. In taluni casi
sia l’uovo interno che quello esterno presentano la stessa anomalia. Le uova
doppie si possono raggruppare in 4 tipi:
·
Uovo
completo dentro a un uovo completo: č un caso piuttosto raro. Nel
1945 Romanoff e Hutt![]() si trovarono in presenza di un eccezionale
caso di produzione di uova doppie in serie: nel giro di tre
mesi
una gallina depose 10 uova doppie complete; nessun
uovo doppio fu preceduto, nel giorno antecedente, dalla deposizione di uova;
due uova doppie furono seguite da uova normali nei giorni successivi;
ordinariamente la deposizione delle uova doppie era separata da un intervallo
di due giorni.
si trovarono in presenza di un eccezionale
caso di produzione di uova doppie in serie: nel giro di tre
mesi
una gallina depose 10 uova doppie complete; nessun
uovo doppio fu preceduto, nel giorno antecedente, dalla deposizione di uova;
due uova doppie furono seguite da uova normali nei giorni successivi;
ordinariamente la deposizione delle uova doppie era separata da un intervallo
di due giorni.![]() .
.
·
Uovo
senza tuorlo dentro a un uovo completo: si tratta della situazione
piů frequente. In un caso descritto da Curtis nel 1916
l’uovo
interno presentava una struttura insolitamente complessa: quattro membrane
concentriche separate da strati d’albume alternativamente chiaro e torbido.
·
Uovo
completo dentro a un uovo senza tuorlo: a questo tipo appartiene l’uovo
completo normale fornito di una seconda serie di involucri.
·
Uovo
avitellino in un uovo avitellino: č il caso piů raro. Dovrebbe
corrispondere all'ovum in ovo descritto da Alberto Magno e riferito da
Conrad Gessner a pagina 422![]() di Historia animalium (1555): "Io ho visto un uovo completamente
sferico ricoperto da due gusci, uno dentro all'altro, con dell'albume acquoso
poco denso che si trovava tra i due gusci e senza tuorlo, e con anche un
secondo albume dentro al guscio piů interno." - Vidi ego ovum prorsus
sphaericum, duabus testis intectum, una intra alteram, cum albumine aquoso
tenui inter utranque absque vitello, et altero etiam albumine intra interiorem
testam.
di Historia animalium (1555): "Io ho visto un uovo completamente
sferico ricoperto da due gusci, uno dentro all'altro, con dell'albume acquoso
poco denso che si trovava tra i due gusci e senza tuorlo, e con anche un
secondo albume dentro al guscio piů interno." - Vidi ego ovum prorsus
sphaericum, duabus testis intectum, una intra alteram, cum albumine aquoso
tenui inter utranque absque vitello, et altero etiam albumine intra interiorem
testam.
Oltre a questi tipi, esistono
casi piů rari e difficilmente classificabili: uova che ne racchiudono altre
due, uovo esterno con 2 tuorli, uova provviste di 3
serie
di involucri, uovo interno giacente nel tuorlo dell’uovo esterno.
Alla gallina domestica spetta il primato di produrre vari
tipi di uova doppie, anche se non mancano esempi in altri uccelli: gallina
della giungla, anatra, oca, tacchina, struzzo. Probabilmente questo fenomeno
puň essere dovuto a una risalita dell’uovo verso la porzione prossimale
dell’ovidutto in seguito a un movimento antiperistaltico.
Lo
afferma anche Frank Lillie in The development of the chick - An introduction
to embryology (1919): "Le uova incluse sono
dovute ad anomale condizioni dell'ovidutto, oppure ad anomalie sia ovariche
che dell'ovidutto. Supponendo che la normale peristalsi dell'ovidutto č
invertita quando un uovo completamente formato vi č presente, l'uovo verrebbe
sospinto nell'ovidutto a una distanza piů o meno grande e vi incontrerebbe un
secondo uovo. Se la peristalsi torna a essere normale, ambedue vengono
sospinti nell'utero e quindi racchiusi in un guscio comune."
Tuttavia
non si puň escludere che a seconda dei casi l'eziopatogenesi
sia piů complessa, come si puň vedere in chiusura del resoconto del Dr
Comellini![]() contenuto nella voce del lessico dedicata all'ovum in ovo
contenuto nella voce del lessico dedicata all'ovum in ovo![]() .
.
|
sommario |
top |
avanti |
|
|
|