Vol. 3° - XII.
Pelle e strutture correlate
|
|
1. cute
1.1. Vesciche al petto
|
Vesciche
al petto |
Poligenico
Gruppo di associazione sconosciuto
Quest’anomalia č comune nei
broilers, sui quali incide in percentuale diversa a seconda del peso, del
sesso e dell’impiumamento del petto; come in quest’ultima
caratteristica, anche per le vesciche sono implicati fattori di tipo
genetico, e verosimilmente siamo di fronte a un tratto poligenico.
1.2. Ittiosi con disidratazione
|
dehy
- dehydrated ichthyosis |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Ittiosi
deriva dal greco ichthýs, che
significa pesce, e si tratta di una discheratosi della cute che appare secca
e finemente desquamante. Alla schiusa i pulcini hanno un piumino che sembra
fil di ferro e sulla schiena si trovano chiazze di cute bianca, ispessita e
desquamante. L’epidermide č quattro volte piů spessa della norma, e nel
giro di una settimana la desquamazione si estende ai tarsi, al ventre, al
becco e agli occhi. Con la perdita delle dita si instaura una zoppia. Quest’anomalia
della cute persiste sino all’etŕ adulta. I soggetti si presentano allora
ricoperti di chiazze ispessite e desquamanti, con una sopravvivenza che non
supera il 40%.
Studiando lo sviluppo
embrionale, si č potuto mettere in evidenza che le prime modificazioni
istologiche compaiono al 19° giorno e che a livello della cute la
moltiplicazione cellulare č da 2 a 3 volte piů intensa rispetto all’embrione
normale. Essendo questo tratto derivato da un ceppo dotato di un difetto
genetico denominato deidratato,
completamente letale, č stata conservata questa denominazione per il
simbolo del gene.
1.3. Sclerodermia
|
sd
- scleroderma |
Autosomico
recessivo, forse associato a geni modificatori
Gruppo di associazione sconosciuto
La
sclerodermia, o sclerosi sistemica progressiva, č una malattia su base
autoimmune, dovuta cioč al fatto che l’organismo aggredisce se stesso
producendo anticorpi diretti contro i normali costituenti tissutali. Ne deriva un processo
infiammatorio cui fa seguito un indurimento della pelle e degli organi per aumento
del connettivo fibroso che va a colmare le lacune in cui si č svolta la
flogosi. In altre parole, dove si č verificata l’infiammazione si
instaura un processo riparativo cicatriziale. Questa mutazione del pollo č
estremamente utile agli studiosi impegnati nel trovare una soluzione all’identico
problema umano.
Quando si ebbero le prime descrizioni di questa malattia si parlava di un fenotipo autosdoppiante, in quanto la cresta dei pulcini, all’etŕ di 2-3 settimane, va incontro a ipertrofia per poi cadere in necrosi parziale. Il risultato finale č una cresta per cosě dire doppia. Durante le prime 6 settimane, oltre a questa progressiva involuzione della cresta ipertrofica, si instaura una fibrosi del derma e una poliartrite distale. Circa il 40% dei soggetti muore entro i 2 mesi a causa di infezioni sovrapposte che si impiantano a livello della cresta e delle dita. I pulcini piů longevi sviluppano una rigiditŕ dell’esofago dovuta a fibrosi, nonché un’infiltrazione mononucleare del cuore e dei reni associata a chiusura dei piccoli e medi vasi sanguigni. Gli esami ematologici mettono in evidenza il fattore reumatoide, anticorpi antinucleo e anticollagene.
2. capo
2.1. Ciuffetto alle orecchie
|
Et
- ear tufts |
Autosomico
dominante
Gruppo di associazione sconosciuto
Abbiamo
giŕ accennato a questa caratteristica nella sezione storica
di questo trattato, quando abbiamo discusso il problema dell’origine del
pollo in Sudamerica (vol.I
- VIII.12 - IX). L’Araucana č unica sotto molteplici aspetti, non solo
per le uova blu, ma anche per il caratteristico ciuffetto alle orecchie,
costituito da un’appendice cutanea coperta di piume che si proietta ai
lati del capo e che č posto in vicinanza del meato uditivo. L’Araucana č
stata ammessa solo di recente nello Standard Inglese, ed esistono delle
differenze tra lo standard Inglese, Tedesco e Americano, per cui i diversi
allevatori hanno preferenze personali circa la forma del ciuffetto.
La natura si sbizzarrisce non solo nel determinare un
aspetto piů o meno piacevole di questo fenotipo, ma anche nell’offrirci
variazioni estreme. Infatti il ciuffetto puň essere sia mono che
bilaterale, puň variare nelle dimensioni, nel modo di proiettarsi del
peduncolo cutaneo, nonché riguardo al punto di origine del peduncolo.
L’espressione di questo gene č pleiotropica, in quanto
determina un rimaneggiamento strutturale dell’orecchio.
Il meato acustico esterno ha una forma molto irregolare e il condotto
uditivo č di lunghezza ridotta oppure č completamente assente. Studi sull’embrione
hanno messo in evidenza che le anomalie anatomiche derivano da una fusione
incompleta dell’arco ioideo con l’arco mandibolare,
e, per questa mancata fusione, il meato uditivo puň variare da un largo
buco a livello della gola fino a una stretta fessura, responsabili del
diverso sviluppo del ciuffetto.
Ralph G.Somes Jr. in The Journal of Heredity (1978)
scriveva cosě: “Attualmente l’Araucana
č una razza morfologicamente molto eterogenea e il suo standard non č
universalmente accettato. L’unico tratto
comune a tutte le varietŕ, e che distingue questa razza dalle altre, č il colore blu delle uova.
Le altre caratteristiche inabituali che le vengono associate sono la
mancanza di coda e il ciuffetto alle orecchie. La presenza o l’assenza di
una o di ambedue queste caratteristiche č apparentemente opzionale per il make-up della razza. Il ciuffetto alle orecchie č espressione dello
stato eterozigote di un singolo gene dominante autosomico. Da notare che, se
č allo stato omozigote, agisce in senso letale prenatale in quasi tutti i
soggetti. Se eterozigote pare si comporti come letale nella fase prenatale
in circa il 20-42%.”
Solo raramente un soggetto con genotipo Et/Et
riesce a sfuggire agli effetti letali del gene. Gli embrioni di questo tipo
muoiono per lo piů tra il 17° e il 19° giorno, e solo pochissimi riescono
a uscire dal guscio, per poi morire in capo a una settimana. Un corollario
pratico per l’allevatore: evitare di accoppiare due soggetti col ciuffetto
se si vuole avere una percentuale di schiusa accettabile.
3. CRESTA

Gallo
con cresta semplice – 1882/1914
xilografia dell'artista inglese Allen William Seaby (1867-1953)
Testo ripreso e adattato dal Giornale degli Allevatori - 1914
La cresta, oggetto di
particolare attenzione sia da parte dell’allevatore che del giudice, ha un
indubbio valore estetico. Ma essa non si limita ad appagare un occhio alla
ricerca del bello, in quanto č un’incontestabile
spia dello stato di salute del soggetto. Per non parlare di coloro che
vorrebbero indossarla come cimiero durante i sollazzi d’alcova, nella
speranza di emulare le doti amatorie
del gallo. Un gallo che ne sia ben dotato, avrebbe anche un’altra
caratteristica, non troppo gradita all’Uomo dei nostri tempi: un’elevata
capacitŕ fecondante.
I soggetti malaticci mostrano una cresta pallida, in certi
casi nerastra, talora coperta di chiazze bianche. Di pari passo vanno i
bargigli, indicando talora un’anemia o una cattiva nutrizione, cui si puň
porre rimedio con una razione generosa e con un tonico ferruginoso, nonché
con una buona ventilazione del pollaio e un supplemento d’aria ossigenata.
Questi provvedimenti danno presto un miglioramento sensibile.
Le malattie del fegato, o la cattiva digestione, fanno
cambiare il colore della cresta da rosso a porpora. Generalmente la causa
risiede nella somministrazione di cibi troppo ricchi di amido e nella mancanza
d’esercizio. Una dieta esclusiva in frumento č un buon rimedio, ed č bene
tenere a mente che i volatili crescono meglio se si dŕ loro solo un pastone
al giorno e con moderazione.
Un poco di sale inglese nell’acqua renderŕ buoni
servigi, come pure la verdura (incluse le cime di cipolla) e un’abbondanza
di grit (la sabbia tagliente). La
mancanza di grit e le creste malate
si danno la mano, e se il ventriglio, che č la macina del pollo, manca di grit,
il fegato e gli organi digestivi peggiorano in breve tempo. In molte covate la
tendenza all’insufficienza epatica comincia dal giorno della nascita,
perché i pulcini sono stati nutriti troppo presto con cibi molli invece che
con grit fine, il quale dovrebbe
essere il primo pasto dei neonati.
Le creste che danno segni di scorbuto per mancanza di
vitamina C, sono spiacevoli alla vista, mentre il favus, o cresta bianca
[1]
,
č molto distruttivo nei pollai. Essendo molto contagioso, si sparge molto
rapidamente. Trattamento ideale č un po’ di iodio, non sotto forma di
tintura, che č irritante, ma uno iodio lavabile, tipo Betadine.
Le creste gelate si possono prevenire sfregandole di
quando in quando con vaselina, cui č stata aggiunta un po’ di glicerina e
di trementina. Č senza dubbio che il gelo attacca indirettamente l’ovaio
attraverso le creste: per questa ragione le galline con piccola cresta sono le
migliori fetatrici invernali.
Durante la muta, spesso la cresta si ritira a metŕ della
sua precedente grandezza. Quando il nuovo piumaggio č completo, e il vigore
ritorna, la cresta riprende le dimensioni primitive per il normale flusso di
sangue.
La cresta con larga base d’impianto č una buona prova
di feconditŕ e vigore nel gallo, visto che si ottengono raramente pulcini
forti e vigorosi da un gruppo ove il gallo ha la cresta piccola.
I galletti seguono il padre e le pollastre la madre nella
forma della cresta, secondo quanto affermano gli scrittori. Ma l’esperienza
degli allevatori dice che la gallina ha piů influenza del gallo.
3.1. Cresta semplice, presenza di cresta
|
Bd+
- allele of bd |
Autosomico
dominante? Poligenico?
Gruppo di associazione sconosciuto
Qualunque
razza dotata di cresta, che non sia quindi la Breda, possiede questo allele.
Il gene della cresta semplice
č indispensabile affinché possa svolgersi l’azione degli altri geni attivi
sulla cresta. Per comoditŕ ho adottato il simbolo Bd+
suggerito da Ab-der-Halden.
La cresta semplice č quella di tipo selvatico,
dentellata, come dimostrano numerosi studi di genetica, ed č posseduta da tre
specie di Galli della giungla: gallus,
sonnerati, lafayettei. Č costituita da una lamina singola che decorre per
tutta la lunghezza della testa, dotata di un numero variabile di dentelli.
Anatomicamente č composta da limbo, denti e lobo.![]()
Le dimensioni della cresta semplice variano da una razza
all’altra, ma č abbastanza stabile nel contesto di ogni singola razza, per
cui si puň pensare a uno stretto controllo genetico. Molte razze a cresta
semplice, specialmente le giganti, hanno una cresta relativamente piccola; le
razze mediterranee, come altre razze, sono dotate di una cresta
proporzionalmente piů grande, talora tanto voluminosa da essere pendula.
Axelsson (1933) ha quantificato queste
differenze, riportando per la Rhode Island una superficie di soli 389 mm˛,
mentre nella Livorno raggiunge 1.321 mm˛. Esistono altri fattori, oltre a
quelli genetici, in grado di influenzare le dimensioni della cresta semplice.
Innanzitutto, credo che qualsiasi allevatore sia conscio del fatto che un
allevamento intensivo č in grado di aumentarne il volume, e ciň dovrebbe
rientrare tra le espressioni indicative di buona salute e di congrua
alimentazione.
Domm, nel 1930, ha fatto notare come le dimensioni sia
della cresta che dei bargigli vengano influenzate in proporzione inversa alla
quantitŕ di luce che il soggetto riceve. Egli ha ipotizzato che il pollo
utilizza la dose necessaria di raggi luminosi solo mediante cresta e bargigli,
che si accrescono per compensare una luce solare scarsa. Non so come possa
essere valida questa supposizione, visto che il Bankiva, che vive nel folto della foresta, ha una cresta
proporzionalmente piccola, mentre le razze mediterranee, inondate di sole,
presentano creste spesso spropositate, quasi antiestetiche, nonché bargigli
tanto lunghi da ricordare lo scroto di un toro.
Nel 1943 Lamoreux condusse una serie d’esperimenti coi
quali fu in grado di dimostrare che aumentando la temperatura da 14°C a 33°C viene triplicata la dimensione della cresta semplice. Pertanto, se un
allevatore desidera una cresta dalle proporzioni ideali, dovrebbe accertarsi
che le condizioni d’allevamento siano corrette. Se vogliamo abbozzare un’interpretazione
finalistica, nell’aumento della superficie della cresta potremmo scorgere
una piů rapida termoregolazione.
Lo standard richiede una cresta
semplice a 5 punte, ma sono frequenti variazioni in piů o in meno. A tale
proposito, si ritiene siano implicati fattori multipli. Inoltre, il locus He+, che modifica la cresta a rosa,
č in grado di influenzare il numero di punte della cresta semplice.
3.2. Assenza di cresta
|
bd
- Breda comb |
Autosomico
recessivo
Gruppo di associazione sconosciuto
La Breda ha preso il nome dall’omonima
cittŕ olandese![]() ed č considerata senza cresta: le femmine ne sono
completamente prive
ed č considerata senza cresta: le femmine ne sono
completamente prive ![]() , i maschi presentano solo due
piccole papille disposte ai lati della linea centrale
, i maschi presentano solo due
piccole papille disposte ai lati della linea centrale ![]() .
In base alle modalitŕ di trasmissione, Bateson e Punnett (1908)
hanno
dedotto che l’allele dominante Bd+
č necessario alla formazione di una cresta. Contemporaneamente dimostrarono
che la Breda č dotata di due alleli D,
responsabili delle due escrescenze nel maschio e delle ampie narici cavernose
caratteristiche di questa razza.
.
In base alle modalitŕ di trasmissione, Bateson e Punnett (1908)
hanno
dedotto che l’allele dominante Bd+
č necessario alla formazione di una cresta. Contemporaneamente dimostrarono
che la Breda č dotata di due alleli D,
responsabili delle due escrescenze nel maschio e delle ampie narici cavernose
caratteristiche di questa razza.
Incrociando un pollo Breda con un soggetto a cresta
semplice potremmo rimanere sorpresi dalla nascita di soggetti con cresta a
rosa: evidentemente il gene R non
puň esplicare la sua azione nel genitore Breda, poiché non č dotato del
gene Bd+ responsabile della cresta.
Una delle migliori descrizioni della Cresta da Breda č dovuta
a Charles-Émile Jacque![]() (Parigi 1813-1894) che cosě scriveva nel 1858 in Le Poulailler:
(Parigi 1813-1894) che cosě scriveva nel 1858 in Le Poulailler:
“La testa
della Breda ha un aspetto del tutto particolare a causa della forma della
cresta, che determina una cavitŕ piuttosto che una prominenza, e dŕ al becco
una depressione caratteristica. Quest’assenza di cresta č tanto piů
interessante in quanto forma, coi bargigli di una bella lunghezza, un
contrasto che non si nota in alcun’altra razza. Deve avere la forma di una
piccola tazza ovale a bordi arrotondati e un po’ sporgenti. Collocata alla
base del becco, sovrasta le narici secondo l’asse della testa, ed č lunga
non piů di 1,5 cm per 1 cm di larghezza. Ha colore nerastro ed č leggermente
corneificata. La cresta, che in alcuni soggetti č trasversale e presenta
delle piccole depressioni sui bordi esterni, non indica sempre una
degenerazione o una mescolanza di sangue, ma questi caratteri ne sono spesso
la conseguenza. Perciň bisogna sempre preferire la cresta ben formata in modo
regolare a foggia di piccola tazza ovale dai bordi uniti.”
Non parlate di Breda a un
Olandese se volete intendere il pollo e non la cittŕ. Infatti, in Olanda, la
razza Breda č denominata Kraaikoppen,
al plurale. Al singolare, cioč kraaikop,
significa testa di cornacchia.

Le Poulailler - 1858
3.3. Cresta a rosa
|
R
- rose comb |
Autosomico
dominante
Gruppo di associazione I - cromosoma 2
Assommano
a piů di una quindicina le razze e le varietŕ che posseggono questo tipo di
cresta, caratterizzata dal fatto di essere larga, quasi appiattita all’apice,
ricoperta da papille piccole e regolari, con la parte terminale posteriore
appuntita, detta anche sperone. Nei
maschi le papille sono piů evidenti che nelle femmine, e la cresta a rosa
delle razze mediterranee č generalmente piů robusta
che nelle altre razze. Fondamentalmente č dovuta all’azione di R
su un genotipo Bd+, per cui si ha la trasformazione di una cresta
semplice, a limbo stretto, in una cresta che, anteriormente, presenta un limbo
molto allargato.
Lunghezza e larghezza della cresta, nonché portamento
dello sperone, dipendono da ciascuna
razza. Per lo piů lo sperone si proietta orizzontalmente, oppure si dirige
lievemente verso l’alto. Nella Barbuta d’Anversa ![]() ,
nella Rhode Island, nella Wyandotte
,
nella Rhode Island, nella Wyandotte ![]() ,
l’intera cresta possiede una curvatura che contorna il cranio, seguendone la
convessitŕ.
,
l’intera cresta possiede una curvatura che contorna il cranio, seguendone la
convessitŕ.
Fu questo gene a dimostrare per primo una piena
rispondenza alle leggi mendeliane. Alle affermazioni di Bateson risalenti al
1902 e che davano per certa un’ereditarietŕ autosomica dominante,
seguirono molteplici verifiche che non apportarono nulla di nuovo. L’unica
indagine a dare un ulteriore contributo allo studio del gene R fu quella di Mérat del 1963, con la quale fu possibile chiarire
che i maschi omozigoti R/R,
anche se appartenenti a razze differenti, sono dotati di scarsa fertilitŕ. I maschi con genotipi R/r+
e r+/r+
hanno una fertilitŕ normale, e tutti e tre i genotipi non compromettono
minimamente le femmine. Innanzitutto, i galli omozigoti hanno una minore
frequenza d’accoppiamento, e la fertilitŕ, dopo un’inseminazione singola,
č di durata piů breve, di circa la metŕ. La motilitŕ degli spermatozoi,
dopo immagazzinamento, č inferiore, e questi gameti differiscono per certe
attivitŕ enzimatiche. Non č ancora del tutto chiarito l’intimo meccanismo
per cui i maschi R/R sono subfertili.
Si puň solo affermare che la subfertilitŕ non č dovuta al genotipo degli
spermatozoi, quanto invece a un effetto deleterio sui gameti da parte dell’organismo
dotato di tale genotipo. Il locus He+,
che modifica il fenotipo della cresta a rosa, puň associarsi a una
riduzione della fertilitŕ maschile.
Gli spermatozoi dei maschi R/R sono dotati di fertilitŕ, ma non sono in grado di competere con
quelli dei galli R/r+.
Questo postulato č dimostrato dalla persistenza della cresta semplice in
quelle popolazioni in cui si effettua un accoppiamento in massa.
Come abbiamo accennato, esistono alcune variazioni a
carico della conformazione della cresta a rosa, probabilmente tutte su base
genetica. Cosě, Punnett propose la
probabile dominanza della punta rivolta verso il basso della Wyandotte
rispetto al fenotipo dell’Amburgo ![]() ;
la stessa dominanza fu dimostrata da Somes tra la cresta della Wyandotte e
quella della Nana di Giava
;
la stessa dominanza fu dimostrata da Somes tra la cresta della Wyandotte e
quella della Nana di Giava ![]() .
.
Spesso, nella Wyandotte, lo standard della cresta a rosa viene disatteso, nonostante reciti cosě:
"La cresta a rosa deve essere
uniformemente e fermamente adesa alla testa, deve possedere una larghezza e un’altezza
media, nella parte anteriore deve essere bassa e squadrata, per affusolarsi
gradualmente man mano che si dirige posteriormente, deve terminare con una
punta ben definita che deve seguire la curvatura del collo senza nessuna
tendenza a dirigersi verso l’alto."
I
difetti ricadono in due categorie principali:
1 - corpo centrale della cresta di dimensioni eccessive e
troppo circolare, nel qual caso č infrequente la presenza di una corretta
superficie dotata di punti piccoli e arrotondati
2 - assenza assoluta di punteggiatura, per cui la
struttura risulta liscia, anche se si avvicina maggiormente alla forma ideale.
Cavalie e Mérat (1965) hanno
scoperto un gene dominante che spiega le differenze osservabili, il gene
della cresta rugosa, hérissée
in francese, contrapposto alla cresta liscia selvatica, senz’altro
responsabile anche nella rugosissima cresta della Redcap ![]() .
.
Altro problema č quello
inerente la deviazione
laterale dello sperone: l’incrocio stretto fra soggetti con tale
fenotipo ha dimostrato che la sua trasmissione non avviene in modo regolare.
Circa Ľ della progenie ha uno sperone deviato, Ľ ha una punta che si esprime
imperfettamente, il 50% č normale. Quando i soggetti con punta abortiva
vengono incrociati tra loro, circa il 62% dei discendenti č dotato dello
stesso tipo di sperone, e il 38% ha uno sperone normale.
3.4. Cresta rugosa · Cresta liscia
|
He+
· hel
- hérissée |
Alleli
di un locus autosomico
Gruppo di associazione sconosciuto
Come
abbiamo giŕ accennato, furono Cavalie e Mérat a proporre questi due alleli,
il cui acronimo deriva dal francese hérissée,
rugosa. Nella cresta a rosa l’allele dominante He+
causa una superficie rugosa giŕ riconoscibile nei pulcini di un giorno,
caratterizzati da una cresta la cui superficie ha un aspetto granuloso, mentre
il soggetto adulto possiede una cresta voluminosa dotata di spuntoni piů alti
e piů numerosi di quelli causati dall’altro allele. Quindi questo allele,
combinato con R, dŕ le creste
larghe e spinose di Nana di Giava ![]() ,
Sebright
,
Sebright ![]() , Redcap
, Redcap ![]() ,
Charollaise
[2]
.
,
Charollaise
[2]
.
Sempre nella cresta a rosa, l’allele recessivo hel
causa
una superficie piů liscia sia nel pulcino che nell’adulto. La sua
penetranza č completa nelle femmine, ma non accade cosě nei maschi, nei
quali pare abbiano influenza alcuni geni modificatori. Combinato con R, dŕ le creste poco rugose della Wyandotte.
I due fenotipi possono essere meglio differenziati nel
pulcino di quanto non accada nell’adulto. Nella cresta semplice ambedue questi alleli sono in grado di
influenzare il numero delle punte,
e i soggetti omozigoti hel/hel
hanno un minor numero di punte rispetto ai soggetti portatori dell’altro
allele dominante. Quindi, nel primo caso, si ha una riduzione del numero dei
denti, che sono anche ben spaziati, come in una bella Nagasaki. Nel secondo
caso, i denti sono piů numerosi e relativamente poco distanziati, elevati,
imprimendo una morfologia a rastrello.
Č interessante il fatto che, in galli a cresta semplice rinchiusi al buio,
la cresta aumenta di dimensioni e rapidamente cade da un lato, ma soprattutto
si notano delle spaziature tra le granulazioni, spaziature la cui superficie
č liscia. Quindi, quando una cresta giŕ formata va incontro a uno sviluppo
supplementare, il numero delle granulazioni non aumenta.
Anche se non sono state effettuate indagini approfondite,
pare verosimile che ambedue gli alleli siano in grado d’influenzare il fenotipo della cresta a noce.
A tale proposito, hel
potrebbe essere responsabile della superficie liscia del cuscinetto che
caratterizza tale tipo di cresta.
Osservazioni di Ab-Der-Halden
inducono pensare che He+ e hel
agiscono solo su Bd+, responsabile della cresta semplice, senza
dipendere in alcun modo da R, da P
oppure da D.
3.5. Cresta a rosa trifida
|
Cresta
a rosa trifida o a spina tripla |
Ereditarietŕ
sconosciuta
La
Barbuta di Watermaal, dotata di fiocco ![]() -
- ![]() , č caratterizzata da una cresta a rosa che termina posteriormente
con 3 punte, la centrale piů lunga delle altre due
, č caratterizzata da una cresta a rosa che termina posteriormente
con 3 punte, la centrale piů lunga delle altre due ![]() -
- ![]() . Č frequente che la cresta mostri la presenza di due soli speroni o di spine malformate.
Finora non si hanno notizie che questo particolare tipo di cresta sia stato
oggetto di studio, ma senza dubbio sono necessari approfondimenti circa l’ereditarietŕ
di questa variante della cresta a rosa. La cresta della Barbuta di Watermaal,
genotipicamente Bd+/Bd+_R/R, puň subire l’azione degli alleli He+
e hel,
ed essere pertanto piů o meno rugosa.
. Č frequente che la cresta mostri la presenza di due soli speroni o di spine malformate.
Finora non si hanno notizie che questo particolare tipo di cresta sia stato
oggetto di studio, ma senza dubbio sono necessari approfondimenti circa l’ereditarietŕ
di questa variante della cresta a rosa. La cresta della Barbuta di Watermaal,
genotipicamente Bd+/Bd+_R/R, puň subire l’azione degli alleli He+
e hel,
ed essere pertanto piů o meno rugosa.
La cresta a rosa della Silky differisce dalle altre in
quanto associata al ciuffo e piuttosto corta, dotata di 2 o 3 punte che si
staccano dalla porzione posteriore. Punnett ha ipotizzato che la
brevitŕ di questa cresta č legata alla presenza del ciuffo, mentre la spina
tripla potrebbe essere dovuta a un altro gene. Studi successivi hanno invece
dimostrato che ambedue le caratteristiche, brevitŕ e punta tripla, possono
essere dovuti alla presenza del ciuffo.
La cresta della Watermaal č simile a quella della Silky e
potrebbe anche risultare dall’interazione tra il gene del fiocco e della
cresta a rosa. Tuttavia Somes ha studiato il fenotipo cresta a rosa trifida in un ceppo senza ciuffo. Incrociando tra loro
soggetti con cresta trifida, ne scaturirono 72 con una punta sola e 155 con
punte multiple. Incrociando soggetti a tre punte con soggetti normali,
nacquero 109 pulcini a punta singola e 91 a punte multiple. Questi dati
starebbero a indicare l’azione di un gene dominante, responsabile della
cresta a rosa a punte multiple.
Che ne pensano i giudici, i
quali non sopportano le tre punte posteriori della Silky? Si tratta proprio di
una cresta abominevole? O č abominevole il voler forzare a ogni costo la
genetica, tollerando o enfatizzando la Watermaal, detronizzando invece la Silky
Triforcuta? Un allevatore olandese, interrogato sulla presenza delle tre
punte nel suo ceppo di Silky, mi ha risposto che in capo a tre anni ciň non
sarebbe piů stato ammesso in Olanda, quindi a partire dal 1997. Pare di essere ai tempi di Hitler! E
spesso, troppo spesso, lo standard ci induce a compiere delle epurazioni che
non hanno nulla da invidiare a quelle perpetrate dal nobile Maestro austriaco!
Credo non sia un’utopia se propongo che un cantuccio dei nostri congressi
mondiali fosse dedicato a questo aspetto dell’allevamento. In altre parole,
gli allevatori
debbono conoscere bene la genetica se non vogliono vedersi costretti a sacrificare al Dio
Standard soggetti meritevoli di vita.
3.6. Cresta a pisello o cresta tripla
|
P
- pea comb |
Autosomico
incompletamente dominante
Gruppo di associazione III - cromosoma 1
Tratto
caratteristico di Brahma, Cornish ![]() ,
Araucana, Cubalaya, Sumatra e altre razze ancora. Talora č denominata cresta
tripla, con 3 linee longitudinali di punti decorrenti dalla fronte verso la
nuca. Si puň riassumere l’azione del gene dicendo che triplica le linee dei dentelli senza triplicare il
limbo. L’allineamento centrale č il piů elevato ed č
quello dotato di dimensioni maggiori.
,
Araucana, Cubalaya, Sumatra e altre razze ancora. Talora č denominata cresta
tripla, con 3 linee longitudinali di punti decorrenti dalla fronte verso la
nuca. Si puň riassumere l’azione del gene dicendo che triplica le linee dei dentelli senza triplicare il
limbo. L’allineamento centrale č il piů elevato ed č
quello dotato di dimensioni maggiori.
La dominanza incompleta non č ben documentabile quando
soggetti con cresta a pisello vengono incrociati con soggetti a cresta
semplice dai bargigli piccoli. Quando vengono accoppiati con razze
mediterranee o con razze dai bargigli imponenti, gli eterozigoti presentano
una lamina centrale ben sviluppata, affiancata da due sporgenze laterali che
rappresentano l’allineamento dei punti esterni.
Munro e Kosin (1940) hanno
dimostrato che a livello del petto i soggetti con cresta a pisello
presentano una striscia di pelle ispessita che
decorre per tutta quanta la lunghezza dello sterno,
associata solo al gene P e forse sua
diretta conseguenza. Quest’area appartiene agli apterili, cioč alle zone
prive delle piume di copertura, e prende il nome di apterilio sternale ![]() .
Questa cute ispessita non causa problemi in una Brahma, dal piumaggio soffice,
mentre puň procurarne alla Cornish, dotata di piumaggio rigido. Il Malese,
secondo le richieste dello standard, trae invece vantaggio da questa
situazione, in quanto si insiste sul fatto che deve essere privo di piume in
sede sternale. Crawford ha voluto verificare questa caratteristica anatomica
anche nei polli con cresta a noce, come il Malese, nel quale si usa parlare di
cresta a fragola. Anche il Malese
presenta la striscia di pelle dura. Il fatto si spiega agevolmente, in quanto
č presente il gene P nel suo patrimonio
genetico. La striscia di cute ispessita puň dimostrarsi utile quando si
vogliono separare i soggetti giovani con cresta a pisello e a noce da quelli
con cresta semplice, in quanto i dati desumibili dalla cresta risultano spesso
insufficienti.
.
Questa cute ispessita non causa problemi in una Brahma, dal piumaggio soffice,
mentre puň procurarne alla Cornish, dotata di piumaggio rigido. Il Malese,
secondo le richieste dello standard, trae invece vantaggio da questa
situazione, in quanto si insiste sul fatto che deve essere privo di piume in
sede sternale. Crawford ha voluto verificare questa caratteristica anatomica
anche nei polli con cresta a noce, come il Malese, nel quale si usa parlare di
cresta a fragola. Anche il Malese
presenta la striscia di pelle dura. Il fatto si spiega agevolmente, in quanto
č presente il gene P nel suo patrimonio
genetico. La striscia di cute ispessita puň dimostrarsi utile quando si
vogliono separare i soggetti giovani con cresta a pisello e a noce da quelli
con cresta semplice, in quanto i dati desumibili dalla cresta risultano spesso
insufficienti.
Numerose ricerche relative a soggetti con genotipo P/p+ e p+/p+, sottoposte a revisione da Mérat e Bordas nel
1979, hanno dimostrato un lieve effetto deprimente sulla velocitŕ di crescita
dovuta al gene P. I lievi effetti
negativi sul peso adulto sono significativi e superano tutti gli altri
parametri presi in esame, mentre i dati sulle uova riguardanti peso, numero,
robustezza del guscio, non vengono intaccati dalla cresta a pisello.
Il drastico rimpicciolimento della cresta e dei bargigli,
responsabili, in condizioni opposte, della dispersione del 15% del calore,
rende ragione della riduzione del 2% del mangime utilizzato dai soggetti con
cresta piccola, in quanto con tali appendici si riduce anche la quantitŕ di
calore dissipato.
Hartmann (1972) ha messo in evidenza la maggior
frequenza di vesciche
al petto nei soggetti con cresta a pisello rispetto a quelli
dotati di cresta semplice. Responsabile pare essere la striscia di pelle
ispessita in tale sede, associata a sua volta a un ridotto impiumamento, come
si puň caratteristicamente notare in diversi Combattenti Malesioidi, dotati
di piumaggio rigido.
3.7. Cresta a noce
|
R
+ P - cresta a noce |
Interazione
di tipo complementare
La
cresta a noce ha la caratteristica di essere bassa, solida e moderatamente
piccola, dotata di una superficie abbastanza variabile ![]() .
Essa risulta dall’azione complementare dei geni della cresta a rosa e della
cresta a pisello. La cresta a noce ha dimensioni minori rispetto alle creste
progenitrici - a rosa e a pisello - e generalmente un’incisura trasversale
separa il terzo posteriore dal rimanente. I pulcini di un giorno, in
corrispondenza del solco, presentano delle piume sottili come un capello.
Questa caratteristica č utile per la loro identificazione, e nell’adulto
questi peluzzi si trasformano in piccole piume.
.
Essa risulta dall’azione complementare dei geni della cresta a rosa e della
cresta a pisello. La cresta a noce ha dimensioni minori rispetto alle creste
progenitrici - a rosa e a pisello - e generalmente un’incisura trasversale
separa il terzo posteriore dal rimanente. I pulcini di un giorno, in
corrispondenza del solco, presentano delle piume sottili come un capello.
Questa caratteristica č utile per la loro identificazione, e nell’adulto
questi peluzzi si trasformano in piccole piume.
Questo tipo di cresta č posseduto da Chantecler,
Kraienköppe
[3]
,
Malese, Orlov, Yokohama, Moroseta. Nella Chantecler
![]() prende il nome di cresta
a cuscino, nel Malese
prende il nome di cresta
a cuscino, nel Malese
![]() viene detta cresta
a fragola, e questa nomenclatura non sta a significare altro che le lievi
differenze esistenti tra le varie morfologie. Nella Silky č richiesta una
cresta di mole ridotta, in quanto deve lasciare spazio al ciuffo.
viene detta cresta
a fragola, e questa nomenclatura non sta a significare altro che le lievi
differenze esistenti tra le varie morfologie. Nella Silky č richiesta una
cresta di mole ridotta, in quanto deve lasciare spazio al ciuffo.
A causa delle dimensioni
ridotte, la cresta a noce č stata introdotta in diverse razze che vivono in
climi rigidi, in quanto č meno suscettibile al congelamento.
3.8. Cresta doppia
|
DV
· DC
- duplex comb |
Autosomico
incompletamente dominante
Gruppo di associazione IV - cromosoma 2
Secondo
Aldrovandi la notizia della presenza di barba associata a una cresta
foggiata - forse - ad anemone sarebbe dovuta a Pausania il Periegeta![]() del II secolo dC
del II secolo dC![]() . Aldrovandi riporta i dati nel brano
seguente, che si trova a pagina 192
. Aldrovandi riporta i dati nel brano
seguente, che si trova a pagina 192![]() del II volume della sua Ornithologia.
Ricordiamo che Tŕnagra
del II volume della sua Ornithologia.
Ricordiamo che Tŕnagra![]() era un’antica
cittŕ della Grecia, in Beozia, nei pressi dell'attuale centro omonimo
situato a 20 km a est di Tebe, situata a sua volta 60 km a nordovest di Atene.
era un’antica
cittŕ della Grecia, in Beozia, nei pressi dell'attuale centro omonimo
situato a 20 km a est di Tebe, situata a sua volta 60 km a nordovest di Atene.
|
Ex
Tanagraeis Gallos potius, quam Gallinas probabant, eorumque bina
erant genera. Alii enim máchimoi,
id est, pugnaces vel proeliares erant, ut Hermolaus vertit: alii
Cossiphi, qui Lydas magnitudine aequabant, quorum Pausanias meminit,
et Corvis colore similes esse tradit (hinc nimirum Cossiphi dicti,
quod Merularum instar atri coloris sint) et barbam, et cristam
habuisse instar anemones [...] |
Dei
polli di Tanagra apprezzavano i galli anziché le galline, e
di essi ne esistevano due razze. Gli uni erano i máchimoi,
cioč come ha tradotto Ermolao
Barbaro erano pugnaci o combattenti; gli altri erano i Cossiphi di
cui ha fatto menzione Pausania, che eguagliavano in grandezza le
galline della Lidia e che dice essere di colore simili ai corvi (per cui
detti appunto Cossiphi [merli] poiché sono di un colore scuro
come quello dei merli) e che avevano sia la barba sia la cresta come un anemone [...] |
Aldrovandi ha preso un bel granchio e lo farebbe prendere anche a noi. Attraverso l’errata interpretazione - in veritŕ non sua - del testo greco di Pausania, induce a pensare che nel II secolo dC in Beozia era presente il gene responsabile di barba e favoriti, e forse anche quello della cresta doppia, con delle chiare implicazioni fuorvianti per coloro che si dedicano alla ricostruzione geografica e storica di una mutazione genetica.
Il
responsabile dell’errata traduzione di Pausania č Abraham Löscher![]() ,
come si puň desumere dal testo di Conrad Gessner, Historia Animalium III (1555), pag. 380
,
come si puň desumere dal testo di Conrad Gessner, Historia Animalium III (1555), pag. 380![]() :
:
Apud Tanagraeos duo genera gallorum sunt, hi machimi, (id est pugnaces, vel praeliares, ut Hermolaus) vocantur, alii cossyphi. Cossyphi magnitudine Lydas gallinas aequant, colore similes corvis (coracino, hinc cossyphi nimirum dicti quod merularum instar atri coloris sint:) barbam et cristam habent instar anemones, (calcaria et apex anemonae [anemones] floris macula<e> modo rubent, Hermol.) Candida item signa exigua in rostro supremo et caudae extremitate, Pausanias in Boeoticis interprete Loeschero.
L’errore in cui grazie a Löscher sia Gessner che Aldrovandi vengono invischiati č molto facile da comprendere. Dobbiamo perň avere la pazienza di imparare il significato di due parole greche: lóphos e kállaion.
Lóphos significa cervice, collo, ma anche ciuffo,
pennacchio, criniera. Questo sostantivo viene tuttavia impiegato per
designare anche quella formazione anatomica carnea che il gallo e la gallina
portano sul capo: la cresta. Infatti Aristotele![]() in Historia animalium 504b cosě si esprime: “Inoltre certi uccelli
presentano una cresta [lóphos] che normalmente consiste di piume
erette; unica eccezione il gallo che ne ha una particolare, formata non
proprio da carne ma di qualcosa non molto dissimile dalla carne.” Il
termine lóphos č di frequente riscontro nella terminologia
scientifica: per esempio Lophophorus, Lophonetta e altri nomi
scientifici di volatili indicano che essi sono dotati di un ciuffo di piume
al capo.
in Historia animalium 504b cosě si esprime: “Inoltre certi uccelli
presentano una cresta [lóphos] che normalmente consiste di piume
erette; unica eccezione il gallo che ne ha una particolare, formata non
proprio da carne ma di qualcosa non molto dissimile dalla carne.” Il
termine lóphos č di frequente riscontro nella terminologia
scientifica: per esempio Lophophorus, Lophonetta e altri nomi
scientifici di volatili indicano che essi sono dotati di un ciuffo di piume
al capo.
Kállaion, singolare e neutro (al plurale kállaia),
viene ugualmente usato da Aristotele per designare la cresta del gallo. Per
lo piů si impiega il plurale kállaia, che assume significato
diverso a seconda del contesto. Kállaia da solo puň significare
cresta, ma puň significare anche bargigli. Il significato di bargigli
diventa inequivocabile se nella frase kállaia č contrapposto oppure
associato a lóphos, dove pertanto lóphos assume il
significato di cresta. C’č di piů. Kállaia, cioč i
bargigli, sono stati equiparati poeticamente alla barba del gallo:
kállaia
dč hoi přgřnes tôn alektryónřn - i bargigli cioč le
barbe dei galli, come
riferisce Ammonio
di Alessandria![]() (VI sec. dC) in De similibus & differentibus dictionibus. Questa trasfigurazione č del tutto logica, se si considera
l’esagerata lunghezza che i bargigli possono talora raggiungere in razze
mediterranee come la Livorno.
(VI sec. dC) in De similibus & differentibus dictionibus. Questa trasfigurazione č del tutto logica, se si considera
l’esagerata lunghezza che i bargigli possono talora raggiungere in razze
mediterranee come la Livorno.
In questa
disamina linguistica possiamo aggiungere che poche pagine piů avanti - a
pagina 196![]() - Aldrovandi dimostra chiaramente di essere
consapevole che sotto il mento di un pollo non c’č una barba, bensě
della pelle membranosa, e cita a suo favore una frase di Columella
- Aldrovandi dimostra chiaramente di essere
consapevole che sotto il mento di un pollo non c’č una barba, bensě
della pelle membranosa, e cita a suo favore una frase di Columella![]() tratta
dal De re rustica VIII,2,9: “Membranosa
illa cutis, quae sub mento, et collo utrinque dependet, palea dicitur: sic
apud Columellam legimus: Paleae ex
rutilo albicantes, quae velut incanae barbae dependent.” E secondo Columella questi bargigli rossi soffusi di bianco
che pendono come le barbe di persone attempate sono un tratto distintivo di
un gallo che č un buon riproduttore. Subito dopo Aldrovandi perde la
pazienza e arriva a tacciare Teodoro Gaza
tratta
dal De re rustica VIII,2,9: “Membranosa
illa cutis, quae sub mento, et collo utrinque dependet, palea dicitur: sic
apud Columellam legimus: Paleae ex
rutilo albicantes, quae velut incanae barbae dependent.” E secondo Columella questi bargigli rossi soffusi di bianco
che pendono come le barbe di persone attempate sono un tratto distintivo di
un gallo che č un buon riproduttore. Subito dopo Aldrovandi perde la
pazienza e arriva a tacciare Teodoro Gaza![]() di avere delle allucinazioni – il che non č del tutto esatto – quando
traduce kállaion di
Aristotele addirittura con cresta: “Hanc membranam, si ita
appellare placet, Aristoteles, vocat
kállaion:
in cuius voce [vocis] traductione Gaza maximopere hallucinatus est,
cristam vertens. Haec enim in vertice erecta est: kállaia
sive paleae utrinque a malis dependent. Videntur autem kállaia dicta ob purpureum, floridumque colorem.”
di avere delle allucinazioni – il che non č del tutto esatto – quando
traduce kállaion di
Aristotele addirittura con cresta: “Hanc membranam, si ita
appellare placet, Aristoteles, vocat
kállaion:
in cuius voce [vocis] traductione Gaza maximopere hallucinatus est,
cristam vertens. Haec enim in vertice erecta est: kállaia
sive paleae utrinque a malis dependent. Videntur autem kállaia dicta ob purpureum, floridumque colorem.”
Insomma, circa i galli di Tanagra Aldrovandi fu indotto in errore da Löscher, ma senza ribellarsi, come invece ha fatto con Gaza.
Pertanto la sequenza etimologica adottata da Löscher circa il significato di kállaia nel testo di Pausania č stata la seguente: kállaia - kállaia dč hoi přgřnes tôn alektryónřn - paleae quae velut incanae barbae dependent - barba. Da cui scaturisce la presenza di barba in una delle due razze di Tanagra, ma solo secondo Löscher.
A pagina 12 di Poultry breeding and genetics di Crawford (1990) possiamo trovare un’interpretazione del testo di Pausania diversa da quella di Löscher, ma che non č meno erronea: “Two breeds were recognized from Tanagra near modern Athens; one was for fighting, and the other was black with crest and wattles ‘like anemones’, which could refer to the rose comb mutant.” Crawford non cita chi ha tradotto il testo greco - peraltro tradotto in modo corretto - e deduce che quella cresta simile a un anemone poteva essere una cresta a rosa, anziché una cresta a coppa, come a mio avviso sarebbe piů logico asserire vista la somiglianza a una coppa del fiore dell’anemone. Dal contesto della frase di Crawford si potrebbe anche evincere che pure i bargigli avessero l’aspetto di una cresta a rosa, ma Crawford non cade in questo tranello.
Abbiamo a disposizione tre interpreti di Pausania che danno torto sia a Löscher che a Crawford: si tratta di D’Arcy Thompson, di W.H.S. Jones e di Ludwig Dindorf.
D’Arcy Thompson in A glossary of Greek birds (1895) alla voce Alektryřn (Gallo) cosě scrive a pag. 39: “At Tanagra, where was a temple <t>o Apollo, there were two breeds, hoí te máchimoi kaě hoi kóssyphoi kaloúmenoi Paus. IX.22.4 - the latter being as black as a crow, with bright red (katŕ anemřnën) comb and wattles and a little white about the beak and tail, leukŕ dč semeîa ou megála epí te ákrři tři rhámphei kaě epě ákras échousi tęs ourâs; this is still a common type of Mediterranean fowls.” Quindi D’Arcy Thompson mette chiaramente in evidenza che quella razza di pollo nera come un corvo aveva cresta e bargigli di un rosso brillante come un anemone.

Pausania il Periegeta - Periegesi della Grecia IX, Beozia, 22.4
W.H.S. Jones cosě ha tradotto il testo di Pausania contenuto nella Periegesi della Grecia IX.22.4: “Here [at Tanagra] there are two breeds of cocks, the fighters and the blackbirds, as they are called. The size of these blackbirds is the same as that of the Lydian birds, but in colour they are like crows [like a crow - kóraki = to a crow], while wattles and comb are very like the anemone. They have small, white markings on the end of the beak and at the end of the tail.”
La traduzione di Jones č estremamente corretta. Giustamente Jones si astiene da ogni preciso paragone cromatico fra anemone e cresta & bargigli, in quanto nel testo di Pausania non compare assolutamente l’aggettivo rosso. La traduzione di Jones suona cosě in italiano: “Qui [a Tanagra] ci sono due razze di galli, i combattenti e i merli, come sono chiamati. Le dimensioni di questi merli sono le stesse di quelle degli uccelli [dei polli, delle galline] della Lidia, ma nel colore essi sono simili a un corvo, mentre i bargigli e la cresta sono molto simili all’anemone; essi posseggono dei piccoli segni bianchi sulla punta del becco e all’estremitŕ della coda.” Attraverso while - mentre - Jones ha messo in risalto la contrapposizione fra il nero del piumaggio e il colore di cresta e bargigli, e ciň corrisponde esattamente al testo greco.
Se non
bastasse, ecco la traduzione altrettanto corretta di Ludwig Dindorf![]() dello stesso passo di Pausania: “Sunt Tanagrae gallinaceorum genera duo,
pugnaces et qui cossyphi (merulae) dicuntur. Hi eadem sunt qua
Lydorum aves magnitudine, colore corvo similes, palearia et crista ad
anemones similitudinem; notas habent candidas in rostri et caudae extrema
parte non utique magnas.”
dello stesso passo di Pausania: “Sunt Tanagrae gallinaceorum genera duo,
pugnaces et qui cossyphi (merulae) dicuntur. Hi eadem sunt qua
Lydorum aves magnitudine, colore corvo similes, palearia et crista ad
anemones similitudinem; notas habent candidas in rostri et caudae extrema
parte non utique magnas.”
Insomma, cosa voleva dire Pausania? Egli voleva semplicemente mettere in risalto come - per una questione di puro contrasto cromatico - cresta e bargigli spiccassero su un piumaggio nero. Inoltre, forse Pausania dava per scontato che il colore piů frequente dell’anemone fosse il rosso brillante. Vediamo se ciň puň corrispondere al vero. Insieme all’anemone analizziamo anche il ranuncolo asiatico, qualora a qualcuno ronzasse per la testa che Pausania abbia voluto parlare magari della cresta a ranuncolo - buttercup - della Siciliana, detta comunemente in italiano cresta a coppa.
L’Anemone coronaria č l’anemone dei giardini per eccellenza; secondo alcuni č originario dell’Oriente ed č presente in Dalmazia, Grecia (quindi senz’altro anche a Tanagra), Turchia (forse Pausania era nato in Lidia), Asia occidentale e si č naturalizzato nella regione mediterranea occidentale.
Il Ranunculus asiaticus, capostipite delle moderne varietŕ coltivate di ranuncoli, č spontaneo a Creta, Cipro, Arcipelago dell’Asia Minore, Siria, Persia, Egitto e Libia. Tutti i ranuncoli contengono succhi spesso dannosi per la presenza di anemonina, posseduta anche dall’anemone, sostanza irritante in grado di causare gravi dermatiti.
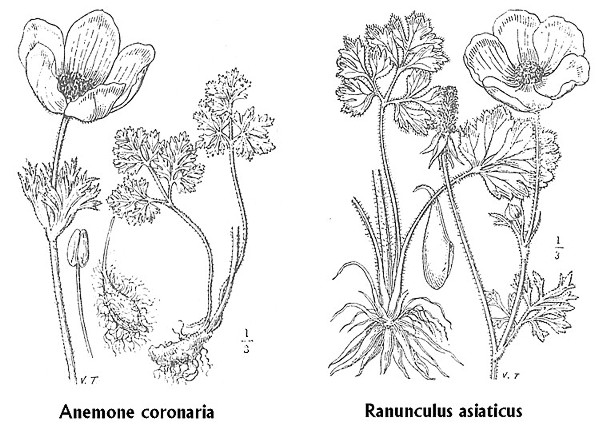
Fig.
XII. 1 – Somiglianza morfologica fra le infiorescenze
di Anemone coronaria
e di Ranunculus asiaticus
Non solo la morfologia, ma anche il colore dei fiori di queste due Ranunculaceae č molto simile. Infatti l’Anemone coronaria varietŕ cyanea presenta fiori azzurri o porporini; nella varietŕ phoenicea o coccinea i fiori sono di un bel rosso cocciniglia. La forma tipica di Ranunculus asiaticus puň avere fiori bianchi, gialli, rossi, porporini o variegati; nella varietŕ sanguineus i fiori possono presentarsi porporini, gialli, aranciati o variegati. Per cui potrebbe essere indifferente da un punto di vista cromatico parlare di cresta ad anemone oppure di cresta a ranuncolo: ambedue i fiori possono essere rossi come abitualmente č rossa una cresta.
Ma
Pausania tra i due fiori ha scelto come termine di paragone l’anemone,
forse perché a lui familiare, o in quanto ai suoi tempi c’era qualcuno
che sbrigativamente chiamava anemone il ranuncolo nonostante il ranuncolo
avesse in greco un nome ben preciso – batráchion – perché vive
volentieri nei luoghi umidi prediletti dalle rane. Sia Dioscoride![]() che Plinio
che Plinio![]() conoscevano quattro specie di ranuncoli, di batráchia, e nel XVI secolo Pierandrea Mattioli
conoscevano quattro specie di ranuncoli, di batráchia, e nel XVI secolo Pierandrea Mattioli![]() arrivň a descriverne sei. Ed č proprio Mattioli a riferire che Pausania
conosceva quel particolare batráchion “acutissimo al gusto” che
“mangiandosi questa herba fa ritirare i nervi, di modo che fa slongar la
bocca, di sorte, che nel morire par propriamente che ridano coloro che se la
mangiano.”
arrivň a descriverne sei. Ed č proprio Mattioli a riferire che Pausania
conosceva quel particolare batráchion “acutissimo al gusto” che
“mangiandosi questa herba fa ritirare i nervi, di modo che fa slongar la
bocca, di sorte, che nel morire par propriamente che ridano coloro che se la
mangiano.”
Ma
Pausania, quando parlando della Sardegna![]() cita questa pianta, non usa la parola batráchion. Egli si limita a
descriverne gli effetti letali e risori, dice che vive nei pressi delle
sorgenti, che č molto simile al sélinon – apium in latino
cita questa pianta, non usa la parola batráchion. Egli si limita a
descriverne gli effetti letali e risori, dice che vive nei pressi delle
sorgenti, che č molto simile al sélinon – apium in latino![]() ,
in italiano detto in passato appioriso – e che ne aveva parlato
Omero. Ma non dice di che colore avesse i fiori, né immagina che si
trattava di un ranuncolo, di un batráchion.
,
in italiano detto in passato appioriso – e che ne aveva parlato
Omero. Ma non dice di che colore avesse i fiori, né immagina che si
trattava di un ranuncolo, di un batráchion.
Invece si
trattava proprio di quel ranuncolo che dalla Sardegna – dove č abbondante
- aveva preso il nome volgare di sardonia o erba sardonica, quell’erba che
messa in bocca fa digrignare i denti provocando il riso sardonico - come
accade anche nel tetano![]() ,
a causa perň dello spasmo muscolare indotto dalla tossina tetanica - erba
alla quale calza perfettamente il nome di Ranunculus sceleratus
appioppatogli da Linneo nel 1753, guarnito perň di fiori piccoli e gialli.
,
a causa perň dello spasmo muscolare indotto dalla tossina tetanica - erba
alla quale calza perfettamente il nome di Ranunculus sceleratus
appioppatogli da Linneo nel 1753, guarnito perň di fiori piccoli e gialli.
Quando
finalmente Pausania usa il termine batráchia non specifica il colore
dei fiori di queste piante. Si limita a paragonare il colore della
capigliatura dei Tritoni![]()
![]() a quello dei batráchia, dei ranuncoli. Ovviamente – come nel caso
dei galli di Tanagra - paragona la chioma dei Tritoni al colore dei fiori
dei ranuncoli che crescono negli acquitrini – tŕ batráchia tŕ en taîs
límnais – e non precisa se i Tritoni avessero le chiome bianche,
gialle, rosse, porporine o variegate.
a quello dei batráchia, dei ranuncoli. Ovviamente – come nel caso
dei galli di Tanagra - paragona la chioma dei Tritoni al colore dei fiori
dei ranuncoli che crescono negli acquitrini – tŕ batráchia tŕ en taîs
límnais – e non precisa se i Tritoni avessero le chiome bianche,
gialle, rosse, porporine o variegate.
In questa
identificazione cromatica non ci viene in soccorso neppure Giglio Gregorio
Giraldi![]() che nel suo Historiae Deorum Gentilium (1548) travisa il testo greco di Pausania
che nel suo Historiae Deorum Gentilium (1548) travisa il testo greco di Pausania![]() presumibilmente identico a quello nelle mani di Ludwig Dindorf: infatti
Giraldi dŕ a batráchia il significato di ranocchie, cosicché i ranuncoli
diventano i colori del dorso delle rane, e i Tritoni anziché avere
dei capelli - che ne so - biondi, si ritrovano ad averli del colore della
schiena delle rane palustri. Per amore della precisione – o della
pedanteria – non possiamo non sottolineare che i Latini per indicare una
piccola rana, un ranocchio, potevano servirsi sia del vocabolo ranula
- utilizzato per esempio da Apuleio – sia di ranunculus, ma
solamente ranunculus indicava anche la pianta di cui stiamo parlando.
Invece se i Greci volevano indicare una piccola rana disponevano del
vocabolo femminile batrachís che troviamo per esempio nella Theriaca
di Nicandro di Colofone
presumibilmente identico a quello nelle mani di Ludwig Dindorf: infatti
Giraldi dŕ a batráchia il significato di ranocchie, cosicché i ranuncoli
diventano i colori del dorso delle rane, e i Tritoni anziché avere
dei capelli - che ne so - biondi, si ritrovano ad averli del colore della
schiena delle rane palustri. Per amore della precisione – o della
pedanteria – non possiamo non sottolineare che i Latini per indicare una
piccola rana, un ranocchio, potevano servirsi sia del vocabolo ranula
- utilizzato per esempio da Apuleio – sia di ranunculus, ma
solamente ranunculus indicava anche la pianta di cui stiamo parlando.
Invece se i Greci volevano indicare una piccola rana disponevano del
vocabolo femminile batrachís che troviamo per esempio nella Theriaca
di Nicandro di Colofone![]() ,
mentre con batráchion esprimevano il ranunculus, la pianta
amante dei luoghi umidi. Ce lo conferma anche Conrad Gessner nel suo Lexicon
Graecolatinum (1537).
,
mentre con batráchion esprimevano il ranunculus, la pianta
amante dei luoghi umidi. Ce lo conferma anche Conrad Gessner nel suo Lexicon
Graecolatinum (1537).

I batráchia di Pausania e di Ludwig August Dindorf
nel suo Pausaniae descriptio Graeciae
IX,21,1
edito a Parigi nel 1845
Né possiamo trovare un conforto cromatico per i capelli dei Tritoni nel sito www.perseus.tufts.edu, in quanto la sequenza verbale conduce alla stessa conclusione di Giraldi, che forse fece da caposcuola. Questo sito afferma che i Tritoni di Pausania avevano i capelli verdi come sono verdi le rane che vivono della paludi.
Nel sito
www.perseus.tufts.edu č presente lo stesso testo greco edito da Ludwig
Dindorf nel 1845. Questo sito riferisce da A Latin Dictionary
di Lewis-Short che ranunculus č sě una piccola rana, č anche un
girino, ma č pure a medicinal plant, called also batrachion, e che ranunculus
ha come sinonimo greco sardanios (che ovviamente non č altro che quel batráchion
oggi chiamato Ranunculus sceleratus o erba sardonica). - Il sito
riporta da A Greek-English Lexicon di Liddell-Scott che
batrachion in prima istanza corrisponde al Ranunculus asiaticus e che
sinonimi di questo batrachion - citato da Ippocrate, nonché da Dioscoride
in II,175 (171 di Mattioli) - sono sia il chrysanthemon (dai fiori gialli)
che il sardanios. Liddell & Scott aggiungono che in Pausania IX,21,1 il
sostantivo batrachion equivale esattamente a bathrachos, cioč rana. Questo
salto mortale – al quale si č sottratto Ludwig Dindorf che forse era
scevro da simili genialitŕ - č stato compiuto da Liddell & Scott al
solo scopo di poter tingere di verde anziché di giallo i capelli dei
Tritoni. Ecco spiegato perché in www.perseus.tufts.edu - sulla scia di
Giraldi – si plagia Pausania e gli si fa dire che i capelli dei Tritoni
hanno il colore delle rane e non dei ranuncoli (che, come le rane, vivono
volentieri nei luoghi paludosi): “I saw another Triton among the
curiosities at Rome, less in size than the one at Tanagra. The Tritons have
the following appearance. On their heads they grow hair like that of marsh
frogs not only in color, but also in the impossibility of separating one
hair from another,” – Ed ecco il testo greco di www.perseus.tufts.edu
identico a quello di Ludwig Dindorf: “Eidon de kai allon Tritôna en tois
Rhômaiôn thaumasi, megethei tou para Tanagraiois apodeonta. Parechontai de
idean hoi Tritônes: echousin epi tęi kephalęi komęn hoia ta batrachia en
tais limnais chroan te kai hoti tôn trichôn ouk an apokrinais mian apo tôn
allôn,” – E cosě, probabilmente sulla scia di Giglio Gregorio Giraldi
– non certo su quella di Ludwig Dindorf - anche William Smith in A
dictionary of Greek and Roman biography and mythology (London. John
Murray. 1873) afferma che i
Tritoni hanno i capelli verdi: “Pausanias (ix. 21. § 1) says: the Tritons have green hair on their head, very fine and hard
scales, breathing organs below their ears, a human nose, a broad mouth, with
the teeth of animals, sea-green eyes, hands rough like the surface of a
shell, and instead of feet, a tail like that of dolphins.” – Č cosě che anche in tempi moderni nascono
i miti, anche i miti relativi ai capelli verdi dei Tritoni, il cui padre,
Poseidone, forse li aveva azzurri, o meglio, il mare gli aveva fatto le mčches
per cui aveva la chioma nerazzurra, come specifica Giraldi nel Syntagma V
del suo Historiae Deorum Gentilium: “Pingebatur Neptunus et ipse
variis modis, nunc pacatus et tranquillus, nunc commotus, ut in primis in
Homero et Vergilio poetis videmus: nunc nudus etiam cum tridente et concha,
quo modo ipse conspexi: nunc in veste caerulea, id est cyanea, ut Phurnutus![]() ait. Lucianus
ait. Lucianus![]() in Sacrificiis, cyaneis capillis et nigris effingit. Sic tamen
a Philostrato
in Sacrificiis, cyaneis capillis et nigris effingit. Sic tamen
a Philostrato![]() in Imaginibus, cum equis scilicet et cetis in mari tranquillo incedere.
[...] Hoc quidem loco Phurnutus ait, propter maris colorem. Unde etiam, ait,
et illum Kyanochaítën appellabant, quod cyaneas et caeruleas iubas
haberet.”– Dopo questa lungaggine non bisogna tralasciare di dire che
Tritone per Ovidio
in Imaginibus, cum equis scilicet et cetis in mari tranquillo incedere.
[...] Hoc quidem loco Phurnutus ait, propter maris colorem. Unde etiam, ait,
et illum Kyanochaítën appellabant, quod cyaneas et caeruleas iubas
haberet.”– Dopo questa lungaggine non bisogna tralasciare di dire che
Tritone per Ovidio![]() era tutto quanto azzurro, laonde le mie chiome bionde se
ne andrebbero a ramengo: “exstantem atque umeros innato murice
tectum|caeruleum Tritona vocat conchaeque sonanti|inspirare iubet fluctusque
et flumina signo” (Metamorfosi I,332-334) Ovviamente da Ovidio non
possiamo arguire se anche tutti gli altri Tritoni successivamente clonati, o
non clonati, fossero completamente azzurri oppure policromi come li aveva
dipinti Pausania. La possibilitŕ dei capelli biondi rimane pertanto aperta!
Quella dei capelli verdi rimane esclusa a priori. E per sempre!
era tutto quanto azzurro, laonde le mie chiome bionde se
ne andrebbero a ramengo: “exstantem atque umeros innato murice
tectum|caeruleum Tritona vocat conchaeque sonanti|inspirare iubet fluctusque
et flumina signo” (Metamorfosi I,332-334) Ovviamente da Ovidio non
possiamo arguire se anche tutti gli altri Tritoni successivamente clonati, o
non clonati, fossero completamente azzurri oppure policromi come li aveva
dipinti Pausania. La possibilitŕ dei capelli biondi rimane pertanto aperta!
Quella dei capelli verdi rimane esclusa a priori. E per sempre!
Oddio!
Liddell&Scott non affermano affatto: batrachion = batrachos
e quindi = verde. Ma lo afferma in modo esplicito William Smith in A
dictionary of Greek and Roman biography and mythology, ovviamente
condizionato dallo stesso condizionamento cromatico che condiziona anche noi
italici quando parliamo di rane (evviva la capacitŕ persuasiva
dell’allitterazione!).
Comunemente,
quando pensiamo alle caratteristiche cromatiche di una rana, il nostro
pensiero corre al verde. Solo in un secondo tempo siamo disposti ad
ammettere tutte le eccezioni cromatiche. Si tratta quindi di un riflesso
condizionato, forse condizionato da molteplici fattori, tra i quali non
bisogna trascurare il fatto che magari la rana che abbiamo piů spesso
osservato era indubbiamente verde, come lo č la rana piů conosciuta in
Italia: la Rana verde comune o Rana esculenta – Rana
esculenta –
cioč commestibile, come indica l’aggettivo latino esculentus,
derivato
dal verbo edo, io
mangio, una rana che č strettamente legata alle acque stagnanti.

Rana verde comune – Rana esculenta
da Guida dei Rettili e degli Anfibi d'Europa di E. N. Arnold and J. A. Burton
edizioni Franco Muzzio & C - in www.ittiofauna.org
Attualmente
in tutto il mondo si conoscono circa 2600 specie viventi di anuri – rane e
rospi - delle quali soltanto 24 si trovano in Europa, e si tratta di specie
abbastanza simili l’una all’altra. Elencheremo le specie presenti
senz’altro in Grecia o nelle immediate vicinanze. Inoltre, sempre per
motivi inerenti alla nostra ricerca, analizzeremo solo il colore del dorso
in quanto č quello che ci permette di pensare istantaneamente al colore
caratteristico di una determinata rana o rospo, e specificheremo il loro
habitat, se cioč vivono di preferenza en taîs
límnais o
se invece utilizzino l’acqua solo durante il periodo riproduttivo. I dati
che seguono sono tratti da Anfibi
e Rettili di
Hans Hvass (1973).
Molto
piů difficile č invece stabilire a quale colore volasse la mente degli
antichi quando pensavano alla rana. Per coloro che abitavano l’Italia č
verosimile il verde della Rana esculenta.
Per
i Greci era possibilissimo il verde, ma non si puň escludere il biondo
scuro grazie alla Rana graeca.

Rana greca – Rana graeca
da Guida dei Rettili e degli Anfibi d'Europa di E. N. Arnold and J. A. Burton
edizioni Franco Muzzio & C - in www.ittiofauna.org
Sta
di fatto che né Aristotele né Eliano fanno il benché minimo accenno al
colore di una qualsivoglia rana, e ne parlano piuttosto frequentemente. Rari
i riscontri cromatici nella Naturalis
historia di
Plinio, che comunque sono riferiti al verde. In XXXII,75 egli parla di una
piccola rana verde dei canneti “Est
parva rana in harundinetis et herbis maxime vivens, muta ac sine voce,
viridis, si forte hauriatur, ventres boum distendens.” – in XXXII,92
parla verosimilmente delle raganella che č verde brillante, ma non ne
specifica il colore “Est rana parva arborem scandens atque ex ea
vociferans; in huius os si quis expuat ipsamque dimittat, tussi liberari
narratur.” - in XXXI,122 parla di una rana verdissima che vive tra
le canne da cui trae il nome greco “Quidam ex ea rana, quam Graeci
calamiten vocant, quoniam inter harundines frutices vivat, minima omnium et
viridissima, sanguinem cineremve fieri iubent,”
Elenchiamo
prima le rane verosimilmente piů note a Pausania e che vivevano nei pressi
di acque stagnanti o di corsi d’acqua. Quindi, per completezza,
elencheremo anche le altre rane nonché i rospi, rimpicciolendo perň i
caratteri tipografici.
Rana
greca – Rana graeca - La
colorazione del dorso č grigia o bruna, di solito con segni sparsi scuri.
Puň essere perň anche rossiccia,
gialla od olivastra. Presente nei Balcani sino alla parte piů
meridionale della Grecia, ma anche in Italia. Vive nei pressi di sorgenti e
corsi d’acqua.
Rana
esculenta – Rana
esculenta - La
colorazione del dorso č per lo piů verde,
soprattutto nei maschi, ma puň essere anche bruna o bluastra. Lungo la
linea dorsale mediana c’č una striscia verde chiara, che talvolta č
invece decisamente gialla. Diffusa ampiamente in Europa sino all’Africa
settentrionale e ad est sino all’Afghanistan, ma assente in Sardegna. Č
una rana essenzialmente acquatica, strettamente legata alle acque stagnanti,
alle famose límnais di Pausania. Č quella cacciata dall’uomo per le sue
carni e venne utilizzata da Luigi Galvani (Bologna 1737-1798) per i suoi
esperimenti che segnarono l'inizio degli studi di elettrofisiologia.
Ululone
dal ventre giallo – Bombina
variegata - Dorso
grigio-bruno o verde oliva, o giallo, con riflessi bronzini. Presente in
tutta l’Europa centrale e meridionale, manca in Gran Bretagna e nella
penisola Iberica. Si trova bene sia in acqua limpida che fangosa.
Ululone
dal ventre di fuoco – Bombina
bombina - Dorso
solitamente
grigio-olivastro con chiazze di colore bruno scuro o verde bottiglia
che nel loro insieme formano un disegno abbastanza simmetrico, che comprende
quasi sempre anche due segni verde chiaro tra le spalle. Presente
soprattutto nell’Europa orientale, con limiti occidentali della zona di
distribuzione che corrono lungo il Weser e il Danubio fino al Mar Nero.
Predilige laghi e stagni.
Rana
arvale – Rana arvalis - Il
colore del dorso varia considerevolmente
dal giallo chiaro al bruno, al grigio, al rossiccio chiaro, sovente
con macchie scure o strisce. Dall’Europa settentrionale e centrale si
estende fino
ai Balcani settentrionali. Vive soprattutto nelle paludi.
Pelobaste
fosco – Pelobastes fuscus -
La colorazione dorsale varia considerevolmente, ma piů sovente č bruno
chiara nel maschio e grigio chiara nella femmina, con segni verde oliva o
bruno-castano misti a piccole chiazze rosse. Specie ampiamente diffusa
nell’Europa centrale ed Orientale, e in Asia, ad eccezione delle zone
montuose. Predilige regioni sabbiose ed entra in acqua solo durante
l’epoca dell’accoppiamento.
Rospo
smeraldino – Bufo viridis -
La colorazione varia considerevolmente: il dorso č grigio chiaro o verde
oliva, con segni irregolari di colore verde erba con margini neri e sovente
con verruche rosse, in particolare sui fianchi. Č specie essenzialmente
dell’Europa orientale e vive in zone costiere sabbiose, quindi non in zone
palustri.
Rospo
comune – Bufo bufo - La
colorazione dorsale – che varia a seconda del sesso, dell’etŕ,
dell’epoca e della localitŕ – va dal nero bruno, al grigio bruno, al
rosso bruno, al rosso olivastro. Gli individui giovani sono per lo piů
giallo bruni o rosso rame, all’incirca come la femmina adulta, mentre i
maschi piů anziani tendono a una tinteggiatura piů uniforme grigiastra od
olivastra. Č l’anuro piů ampiamente distribuito in Europa. Trascorre il
giorno nei giardini, nei campi, nei boschi, anche tra le vegetazione fitta.
Puň essere rintracciato anche nelle cantine. Quindi non vive negli
acquitrini. Solo in aprile le coppie restano insieme notte e giorno e
trascorrono in acqua la maggior parte del tempo.
Rana
dalmatina – Rana dalmatina -
Il colore bruno del dorso puň sfumare nel grigio pallido o nel giallo
torbido. Vive in boschi decidui aperti, litorali assolati e prati. Č
presente dalla Francia ai Balcani, compresa l’Italia, e a est in Asia
Minore e nel Caucaso.
Raganella – Hyla arborea - La colorazione č di solito verde brillante sul dorso, ma a seconda di svariati fattori puň essere verde cupo, gialla, grigia, azzurra, bruna o anche nera. Presente nell’Europa meridionale e centrale, vive in acqua solo in coincidenza del periodo della deposizione delle uova. Nelle altre stagioni si trova per lo piů sugli alberi, fra i cespugli e nei canneti.
Ma i Tritoni vivevano nell’acqua di mare e non di palude, per cui potrebbe essere nel giusto Ovidio quando dice che Tritone era tutto quanto azzurro, con le chiome che ricalcavano quelle del padre Nettuno. Siccome i Tritoni non vivevano negli stagni e nei fiumi, non si vede perché dovessero avere i capelli verdastri anziché azzurri oppure biondi. Anche se Pausania non ce lo dice, possiamo dedurre da Dioscoride - nonché da Galeno - che a quei tempi il colore prevalente del ranuncolo era senz’altro il giallo. Infatti per Dioscoride un tipo di ranuncolo aveva fiori gialli, talora rossi (intendendo verosimilmente il Ranunculus asiaticus), due avevano sempre i fiori gialli (incluso il Ranunculus sceleratus), uno sfoderava fiori candidi come il latte. Quindi ai tempi di Dioscoride e di Pausania a carico dei ranuncoli prevaleva il giallo, ed č verosimile molto assai che per Pausania i Tritoni fossero biondi, cosě come ha inteso che fossero rossi cresta e bargigli quando ha scritto “ma nel colore essi sono come i corvi, mentre i bargigli e la cresta sono molto simili all’anemone”, riferendosi con ogni probabilitŕ al colore rosso cocciniglia dell’Anemone coronaria nella varietŕ phoenicea o coccinea, tralasciando un qualsivoglia batráchion dal fiore rosso, altrimenti avrebbe scritto che cresta e bargigli erano come un batráchion (specificando rosso) anziché come un anemone.
Se vogliamo essere pignoli e pedanti - o precisi per dirla eufemisticamente - non possiamo escludere che Pausania pensasse magari al fiore di quella Ranunculacea, a quell’anemone che nel 99,9% dei casi č rosso, battezzato poi Adonis annuus da Linneo nel 1753 e che nel XVI secolo - per esempio da Mattioli - era ancora chiamato anemone.
Per gli antichi Greci l’anemřnë – in dorico anemřna, talora anemřnion in Dioscoride - era quel fiore che si apre al minimo vento, o che talora cresce nei luoghi ventosi, come indica la sua etimologia da ánemos che significa vento, soffio, come il latino anima. Plinio propendeva per il primo dei due significati: “Flos numquam se aperit nisi vento spirante, unde et nomen accipere.” (XXI,165)
In greco antico non esiste nulla che si chiami adone - ádřnis
- se non un pesce citato da Ateneo (332c) e da Eliano![]() nonché da Plinio
nonché da Plinio![]() .
.
Ádřnis era invece Adone ![]() , quel bellissimo ragazzo del quale si innamorň Venere
e che morě assalito da un enorme cinghiale. In base a un calcolo
percentuale – infatti non ero presente in Libano quando Adone morě - dal
suo sangue sarebbe nato un anemone, che in realtŕ era quel fiore che oggi
grazie a lui chiamiamo Adonis annuus
, quel bellissimo ragazzo del quale si innamorň Venere
e che morě assalito da un enorme cinghiale. In base a un calcolo
percentuale – infatti non ero presente in Libano quando Adone morě - dal
suo sangue sarebbe nato un anemone, che in realtŕ era quel fiore che oggi
grazie a lui chiamiamo Adonis annuus![]()
![]() ,
e ambedue le varietŕ annue, sia la aestivalis o phoeniceus
che la autumnalis o atrorubens, hanno fiori color del sangue.
L’Adonis annuus cresce dal livello del mare fino ad altitudini di
1300 m, č presente in Europa, Caucaso, Africa boreale e, ciň che piů
conta, in Asia centrale e occidentale, quindi anche in Libano dove Adone morě.
,
e ambedue le varietŕ annue, sia la aestivalis o phoeniceus
che la autumnalis o atrorubens, hanno fiori color del sangue.
L’Adonis annuus cresce dal livello del mare fino ad altitudini di
1300 m, č presente in Europa, Caucaso, Africa boreale e, ciň che piů
conta, in Asia centrale e occidentale, quindi anche in Libano dove Adone morě.
Adone: divinitŕ di origine semitica, tipizzata sul prototipo del dio Tammuz (il dio assiro-babilonese della vegetazione, giŕ appartenente al pantheon sumerico come Dumuz) che rappresentava la perenne trasformazione della vegetazione e che annualmente risorgeva sotto lo stimolo di un culto. Adon in cananeo significava signore, presente nell’ebraico biblico Adonai, mio signore, appellativo di Dio nell’Antico Testamento. Introdotto nel mondo greco-romano (la prima testimonianza di Adone la troviamo in Saffo), il dio ha come paredra Afrodite (paredra era la dea che si accompagnava a un dio, che gli sedeva accanto, spesso come moglie, assolvendo gli stessi compiti) e nella nuova versione del mito perde il suo alone divino, trasformandosi in eroe. Secondo la mitologia greca Adone nacque grazie al rapporto incestuoso tra Cinira, re di Cipro e sacerdote di Afrodite, e la figlia Smirna o Mirra che, innamoratasi del padre, si fece passare per una delle mogli mediante un sotterfugio. Quando il padre scoprě l’incesto in cui era caduto, Mirra fu costretta a fuggire, e gli dei per salvarla la tramutarono in una pianta resinosa dall’amaro profumo, che da lei prese il nome. A primavera la corteccia della pianta miracolosamente si aprě e ne uscě il piccolo Adone. Afrodite raccolse il neonato, lo chiuse in un cofano e lo affidň, perché lo allevasse, a Persefone regina dell’Erebo, sposa di Ade il re degli Inferi, che era il regno delle tenebre e la dimora delle anime dei morti. Il bimbo crebbe rapidamente e divenne un bellissimo adolescente. Allora Afrodite si recň nell’Erebo per riprenderlo, ma Persefone non volle restituirlo. Zeus, per conciliare il contrasto tra le due dee, innamorate entrambe del giovane, decise che Adone avrebbe trascorso un terzo dell’anno con Afrodite, un altro terzo con Persefone, lasciandolo libero di disporre per sé del restante tempo. Ma Adone preferiva trascorrere anche questo terzo con Afrodite, suscitando la gelosia di Ares – o Marte – e alla fine di un’estate, mentre era a caccia sui monti del Libano, uno smisurato cinghiale mandato dal geloso Ares assalě e uccise Adone procurandogli una ferita all’inguine. Afrodite accorse per salvarlo, ma non giunse in tempo. - E qui viene il bello. Ripeto che non c’ero sui monti del Libano quando il cinghiale di Ares uccise Adone. Dobbiamo quindi affidarci alle percentuali di probabilitŕ derivanti dalle 19 fonti consultate: cosě risulta vincente la leggenda secondo cui dal sangue di Adone scaturě un anemone (9 fonti), l’anemone nacque dal corpo di Adone - i cui muscoli erano senz’altro rossi come lo era il sangue - (4 fonti), una fonte dice che il fiore scaturě dalle lacrime di Afrodite, un’altra afferma che le rose che si trovavano vicino ad Adone da bianche divennero rosse, una fonte dichiara che Afrodite creň l’anemone dal nulla allo scopo di avere un simbolo dell’amato presso di sé, e infine 3 fonti non citano assolutamente Adone come responsabile della nascita dell’anemone, né di qualsivoglia altro fiore. Alle fonti che depongono per un’emogenesi dell’anemone appartiene anche Ovidio, che cosě scrive in Metamorfosi X, 728: at cruor in florem mutabitur e al verso 735: cum flos de sanguine concolor ortus. Ma una fonte per la somatogenesi del fiore color sangue č rappresentata da Conrad Gessner che alla voce Adonis cosě scrive nel suo Onomasticon propriorum nominum (1564): “Nomen proprium filii Cynarae Cypriorum regis et Myrrhae eius filiae, quem Venus in delitiis habuit formositate praecellentem. [...] Venus miserata post multas lachrymas in florem Adonium, qui est sanguineo colore, eum transformavit.” Quindi, grazie al mito vincente, quando Pausania pensava a un anemone, pensava al colore rosso, in quanto il fiore era sbocciato dal sangue di Adone.
Invece l’Adonis vernalis ![]() , che č perenne anziché annuale, ha
fiori gialli, cresce in climi piů freddi e non č presente in Asia se non
in Siberia. Mattioli aveva individuato 5 diversi anemoni, e quello che
aveva catalogato come Anemone quarta, dai fiori porporini, potrebbe
corrispondere all’Adonis annuus di Linneo.
, che č perenne anziché annuale, ha
fiori gialli, cresce in climi piů freddi e non č presente in Asia se non
in Siberia. Mattioli aveva individuato 5 diversi anemoni, e quello che
aveva catalogato come Anemone quarta, dai fiori porporini, potrebbe
corrispondere all’Adonis annuus di Linneo.
Insomma, grazie al mito, quando Pausania pensava a un anemone, pensava al colore rosso, e magari pensava a quell’anemone il cui fiore era sbocciato dal sangue di Adone: Adonis annuus.

Fig. XII. 2 – sopra: Anemone quarta di Pierandrea Mattioli. Si tratta verosimilmente dell’Adonis annuus di Linneo, detto Adonide o Fiore di Adone, tratto dal Compendium de plantibus omnibus (1571) di Mattioli. – sotto: Adonis annua, da N.L Britton e A. Brown, Illustrated flora of the northern states and Canada. (1913).

Fig. XII. 3 – Gallo con cresta doppia.
Particolare di un mosaico del II secolo dC rinvenuto nel 1801 a Poggio
Mirteto (Rieti) al cui centro si trova una Diana di Efeso![]() con corona turrita da cui scende un velo, tre linee di mammelle sul petto,
corpo trasformato in fusto e piedi nudi. Il mosaico č conservato presso il
Museo Chiaramonti dei Musei Vaticani (da Poultry Farming as described by
the writers of ancient Rome, 1939). - Per ulteriori notizie sul mosaico
vedi Bartolomeo Nogara, I mosaici antichi conservati nei palazzi pontifici,
1910, p. 27, tav. LV.
con corona turrita da cui scende un velo, tre linee di mammelle sul petto,
corpo trasformato in fusto e piedi nudi. Il mosaico č conservato presso il
Museo Chiaramonti dei Musei Vaticani (da Poultry Farming as described by
the writers of ancient Rome, 1939). - Per ulteriori notizie sul mosaico
vedi Bartolomeo Nogara, I mosaici antichi conservati nei palazzi pontifici,
1910, p. 27, tav. LV.
Plinio
conosceva la cresta doppia, raffigurata anche nel mosaico rinvenuto a Poggio
Mirteto e conservato presso il Museo Chiaramonti dei Musei Vaticani. Vediamo
cosa dice Plinio, riportando il brano che abbiamo giŕ avuto modo di
analizzare nel primo volume.
|
Gallinarum
generositas spectatur crista erecta, interim et gemina, pinnis
nigris, ore rubicundo, digitis imparibus, aliquando et super IIII
digitos traverso uno. (X,156) |
La
buona razza delle galline si riconosce dalla cresta eretta, talvolta
anche doppia, dalle piume nere, dalla faccia rossa, dalle dita di
differente lunghezza, e talvolta anche dalla presenza di un dito
disposto obliquamente oltre agli altri quattro. |
Ai
tempi di Aldrovandi la cresta doppia variamente foggiata era molto diffusa,
come ci attestano sia l’abbondante iconografia in bianco e nero contenuta
nella sua Ornithologia, sia le immagini a colori di cui aveva curato
l’esecuzione ![]() .
.

Fig. XII. 4 – Ulisse Aldrovandi - 1600 - Gallina pumilio capite cirrato

Fig. XII. 5 – Ulisse Aldrovandi - 1600 - Gallus et Gallina Patavini

Fig. XII. 6 – Ulisse Aldrovandi - 1600 - Gallus et gallina pedibus pennatis

Fig. XII. 7 – Ulisse Aldrovandi - 1600 - Gallus et Gallina Turcici

Fig. XII. 8 – Ulisse Aldrovandi - 1600 - Gallus et Gallina Persici

Fig. XII. 9 – Ulisse Aldrovandi - 1600 - Gallus crispus

Fig. XII. 10 – Ulisse Aldrovandi - 1600 - Gallina lanigera
Gli alleli del locus D
causano uno sdoppiamento di tutta la cresta o solo di una sua porzione. Nel secondo caso la parte posteriore della cresta si
sdoppia piů frequentemente dell’anteriore ![]() -
- ![]() . Non si č mai osservato lo
sdoppiamento della sola parte anteriore. Quest’azione fondamentale,
associata a quella di geni attivi sulla forma della cresta, determina
fenotipi particolari. Per esempio, partendo da una cresta semplice si
ottiene la cresta a corona o a coppa della Caumont
[4]
e la cresta a foglia di quercia della Houdan
[5]
che somiglia a una farfalla con ali semiaperte disposta
davanti al ciuffo
. Non si č mai osservato lo
sdoppiamento della sola parte anteriore. Quest’azione fondamentale,
associata a quella di geni attivi sulla forma della cresta, determina
fenotipi particolari. Per esempio, partendo da una cresta semplice si
ottiene la cresta a corona o a coppa della Caumont
[4]
e la cresta a foglia di quercia della Houdan
[5]
che somiglia a una farfalla con ali semiaperte disposta
davanti al ciuffo ![]() -
- ![]() -
- ![]()
Gli alleli del locus D
intervengono anche nel determinare il doppio corno di La Flčche
[6]
, Crčvecoeur
[8]
![]() -
- ![]() , Ciuffata di Appenzell
[7]
, Ciuffata di Appenzell
[7]
![]() ,
Polish
[9]
e Sultano. In questi casi all’azione del gene della cresta doppia si associa l’effetto
di qualche gene modificatore, per cui il fenotipo puň variare parecchio sia
per interazione con geni responsabili della forma della cresta, sia per
interazione con geni modificatori non specifici. Infatti esistono geni modificatori i cui alleli impongono una forma
particolare alla cresta sdoppiata da D.
Questa soluzione, anche se incompletamente soddisfacente, sembra essere
plausibile.
,
Polish
[9]
e Sultano. In questi casi all’azione del gene della cresta doppia si associa l’effetto
di qualche gene modificatore, per cui il fenotipo puň variare parecchio sia
per interazione con geni responsabili della forma della cresta, sia per
interazione con geni modificatori non specifici. Infatti esistono geni modificatori i cui alleli impongono una forma
particolare alla cresta sdoppiata da D.
Questa soluzione, anche se incompletamente soddisfacente, sembra essere
plausibile.
La Siciliana - chiamata Sicilian buttercup e talora
solo Buttercup in inglese - ha una cresta a coppa o a
ranuncolo, che in inglese č detto buttercup: si tratta di una
coppa foggiata a corona circolare ![]() -
- ![]() con punti - o dentelli - di media grandezza e regolari, la quale si
appoggia al centro del cranio.
con punti - o dentelli - di media grandezza e regolari, la quale si
appoggia al centro del cranio.
Le femmine di Breda sono completamente senza cresta ![]() ,
mentre, come abbiamo giŕ detto, i rispettivi maschi presentano solo due
piccole papille disposte ai lati della linea centrale
,
mentre, come abbiamo giŕ detto, i rispettivi maschi presentano solo due
piccole papille disposte ai lati della linea centrale ![]() .
In base alle modalitŕ di trasmissione, Bateson e Punnett (1908)
hanno
dedotto che l’allele dominante Bd+
č necessario alla formazione di una cresta e contemporaneamente
dimostrarono che la Breda č dotata di due alleli D,
responsabili delle due escrescenze nel maschio e delle ampie narici
cavernose caratteristiche di questa razza.
.
In base alle modalitŕ di trasmissione, Bateson e Punnett (1908)
hanno
dedotto che l’allele dominante Bd+
č necessario alla formazione di una cresta e contemporaneamente
dimostrarono che la Breda č dotata di due alleli D,
responsabili delle due escrescenze nel maschio e delle ampie narici
cavernose caratteristiche di questa razza.
Infatti una caratteristica comune a tutte le razze con
cresta doppia, eccetto la Siciliana, consiste nelle narici ampie e cavernose. L’ereditarietŕ di questo
tratto ha comportamento recessivo.
Tutti i fenotipi descritti sono dovuti a un unico locus D, e pare possibile l’associazione con un’ernia cerebrale.

Gallus
cornutus - Gallus Indicus
Avium Vivae Icones - 1600
di Adriaen Collaert (Anversa c. 1560 - 1618)
Somes (1986) ha dimostrato che il fenotipo a
cornetti, o a lettera V, e il fenotipo a coppa come nella Siciliana, sono
comandati da due diversi alleli, aggiungendo cosě la lettera v
al primo, e la lettera c al
secondo per significare coppa.
L’allele Dv
č dominante sul suo
gemello, č potente nel duplicare una cresta e ha una penetranza completa.
L’allele
Dc
č
dotato di un’azione sdoppiante piuttosto fiacca e nelle femmine la sua
penetranza si riduce del 25%.
Si puň pertanto desumere che le narici ampie e cavernose si esprimono appieno nel
genotipo Dv/Dv,
ma non in quello eterozigote Dv/d+. L’altro allele si esprime con narici ampie solo
se associato a quello della cresta a V: Dv/Dc.
La cresta a forma di foglia di
quercia non č stata studiata a fondo, ma pare probabile si debba invocare
un terzo allele per il locus D.
L’arrivo del Sultano in Inghilterra - citazione di Lewis Wright (1890)
Il Sultano fu introdotto in
Inghilterra da Miss E. Watts, e in base alla citazione del testo di Miss
Watts da parte di Wright![]() nel suo The illustrated book of Poultry i fatti si svolsero cosě:
nel suo The illustrated book of Poultry i fatti si svolsero cosě:
“Questo pollo mi fu inviato, nel gennaio 1854, da un
amico che vive a Costantinopoli. L’anno prima gli avevo mandato alcuni
esemplari di Cocincina, che gli piacquero moltissimo, e successivamente suo
figlio venne in Inghilterra. Disse che poteva mandarmi dalla Turchia dei polli
che mi sarebbero senz’altro piaciuti. Tutte le interessanti caratteristiche
di questa razza giunsero prima del pollo, rendendomi oltremodo curiosa di
poterle apprezzare dal vero. Approdarono finalmente in gennaio, a bordo di un
piroscafo il cui equipaggio era prevalentemente costituito da Turchi. Il
viaggio era stato lungo e tempestoso, per cui i poveri polli si erano rotolati
nel sudiciume e si erano impiastricciati in un modo mai visto prima. Ripuliti
a dovere, me ne innamorai subito, ma, non volendo correre rischi per la
riproduzione, scrissi subito di inviarmi altri soggetti, soprattutto un gallo,
chiedendo anche qual era il loro nome. L’amico rispose che gli era difficile
trovare dei begli esemplari, che avrebbe cercato di mandarmene altri due o
tre, ma che le sue ricerche sia a Costantinopoli che in altre parti della
Turchia risultavano infruttuose. Non appena avesse trovato un soggetto a
dovere, me lo avrebbe inviato.
“Circa il nome, mi fece sapere che in Turchia questi
polli sono chiamati Serai-Täook
[10]
.
Serai č il Serraglio, cioč il
palazzo del Sultano, e nei Paesi di civiltŕ turca indica la sede di un
governo o la residenza di un principe. Täook
in turco significa pollo[fowl]. Per cui, la traduzione piů semplice č Polli
del Sultano [Sultan’s fowls], nome che presenta il doppio vantaggio di essere il piů
vicino a quello usato nella patria d’origine e di indicarne il luogo di
provenienza.”
“[...] Hanno le dimensioni dei nostri Polish inglesi. Il loro piumaggio č bianco e fluente, hanno sulla testa il ciuffo pieno e compatto della Polish, hanno i favoriti, una bella coda fluente, delle zampe corte ben impiumate e cinque dita ad ogni zampa.”

Lewis Wright - 1838-1905
Quindi i soggetti inviati a Miss Watts erano pentadattili. Terminata la citazione di Miss Watts, Lewis Wright ci tiene a precisare che nel 1890 la pentadattilia del Sultano non era la regola e ci si poteva imbattere in soggetti sforniti del quinto dito e con basette mignon: “Even the prize birds shown in the plate, it will be seen, have little muffling, and are destitute of the fifth toe.” Sul perché dell’incostanza della pentadattilia si veda Pentadattilia europea: celtica o romana? (X,8).
Anche
Darwin afferma che il Sultano č originario della Turchia. Alcuni invece
pensano che si sia formato in Russia, per essere poi allevato a Costantinopoli
(Teodoro Pascal![]() , 1905). Come
vedremo fra poco, č verosimile che Pascal abbia tratto la notizia
dell’origine russa del Sultano dagli scritti di Dürigen, autore da lui
citato in alcune occasioni. Pascal, a differenza di Miss Watts, dŕ un’altra
grafia del nome turco di questa razza. Infatti egli scrive: “Questa razza,
chiamata a Costantinopoli Serai-Taook da serai che vuol dire
serraglio e Taook gallina, proviene dalla Turchia, ma taluni la dicono
formata in Russia e quindi allevata a Costantinopoli.” Sia Pascal che Miss
Watts scrissero del Sultano prima del 1928, data in
cui, per impulso di Kemal Atatürk, fu introdotto l'alfabeto latino con
particolari segni diacritici che sostituě l'alfabeto arabo - e alcuni
caratteri aggiunti dai Persiani - usato appunto fino al 1928. Oggi in turco la gallina si scrive Tavuk, per cui
Täook, Ta-ook e Taook potrebbero essere delle traslitterazioni
dall’alfabeto arabo e dovrebbero equivalere all’attuale Tavuk. Se tavuk č
la gallina, horoz č il gallo, piliç č il pollo e civciv č
il pulcino.
, 1905). Come
vedremo fra poco, č verosimile che Pascal abbia tratto la notizia
dell’origine russa del Sultano dagli scritti di Dürigen, autore da lui
citato in alcune occasioni. Pascal, a differenza di Miss Watts, dŕ un’altra
grafia del nome turco di questa razza. Infatti egli scrive: “Questa razza,
chiamata a Costantinopoli Serai-Taook da serai che vuol dire
serraglio e Taook gallina, proviene dalla Turchia, ma taluni la dicono
formata in Russia e quindi allevata a Costantinopoli.” Sia Pascal che Miss
Watts scrissero del Sultano prima del 1928, data in
cui, per impulso di Kemal Atatürk, fu introdotto l'alfabeto latino con
particolari segni diacritici che sostituě l'alfabeto arabo - e alcuni
caratteri aggiunti dai Persiani - usato appunto fino al 1928. Oggi in turco la gallina si scrive Tavuk, per cui
Täook, Ta-ook e Taook potrebbero essere delle traslitterazioni
dall’alfabeto arabo e dovrebbero equivalere all’attuale Tavuk. Se tavuk č
la gallina, horoz č il gallo, piliç č il pollo e civciv č
il pulcino.

Gallo e gallina di razza Sultano di Teodoro Pascal – 1905. Contrariamente alle caratteristiche indicate dall’American standard of perfection (1993), ai tempi di Teodoro Pascal “il numero delle dita, nella razza tipica, deve ascendere a cinque, ma si ammettono anche individui a quattro dita.” (da Le razze della gallina domestica) Inoltre Pascal, pur affermando che la barba č piuttosto corta, ma spessa e decorata di basette corte e folte, nella sua raffigurazione sia il gallo che la gallina sono imberbi e senza basette, con il maschio che ha dei bargigli da Livornese, e che quindi non sono “piuttosto corti (3 cm)” come descritto nelle note caratteristiche, ma assai esuberanti. Insomma, nel 1905 si accettavano anche le 4 dita, una barba fantasma e dei bargigli rigogliosi.


Sultano imberbe e barbuto di Lewis Wright (1838-1905) – Ambedue le immagini sono dovute al talento artistico di J. W. Ludlow. I soggetti senza barba risalgono al 1872 in quanto erano presenti nella prima edizione del The Illustrated Book of Poultry di Wright e appartenevano a Mr Robert Loft. Invece i soggetti con barba e favoriti non sono databili con certezza, ma sono comunque posteriori al 1872. Anche Wright afferma che questa razza č dotato di 5 dita – come i soggetti giunti a Miss Watts da Costantinopoli – ma che talora aveva solo 4 dita.
Molto
precise sono le parole di Bruno Dürigen, dal quale Pascal ha verosimilmente
tratto la notizia della possibile origine russa del Sultano. Infatti Dürigen
cosě scriveva a pagina 211
del suo Die Geflügelzucht (1921):
54.
Türken. Dieses Haubenhuhn mit Federfüßen und Bart - Gallus dom. barbato·cristatus,
plumipes - erinnert von allen Türken Haubenhühnern am meisten an die mutmaßliche
Stammform, nämlich das in Rußland seit Jahrhunderten heimische federfüßige
Bart-Haubenhuhn (S. 212) [55.
Pawlowa (Russen)].
54. Turchi. Questo pollo ciuffato dotato di piume alle zampe e di barba - Gallus domesticus barbato-cristatus, plumipes [Gallo domestico barbuto-ciuffato, con le piume ai piedi] - ricorda maggiormente, tra tutti i polli ciuffati, quello che con ogni probabilitŕ ne č l'antenato, vale a dire il pollo dotato di zampe piumate, di barba e di ciuffo, da secoli di casa in Russia (pagina 212) [cioč, 55. Pavlov (Russi)]. (traduzione di Stefano Bergamo, 2003)
Č
tuttavia possibile che un pollo, oriundo comunque della Cina, abbia raggiunto
Costantinopoli e la Turchia attraverso una via diversa da quella russa:
infatti nell’XI secolo i Turchi Selgiuchidi penetrarono in Anatolia
dopo aver sconfitto l’esercito dell’imperatore bizantino Romano IV Diogene a Malazgirt (1071). John
Peters (The cock, J. Ameri. Oriental Soc. 33:363-396,
1913![]() ) scrisse: “Tuttavia
probabilmente č stato dalla Cina che il pollo passň ai Turchi e piů tardi
venne portato a occidente con loro, in un momento in cui giŕ da lungo tempo
era stato addomesticato in Occidente.”
) scrisse: “Tuttavia
probabilmente č stato dalla Cina che il pollo passň ai Turchi e piů tardi
venne portato a occidente con loro, in un momento in cui giŕ da lungo tempo
era stato addomesticato in Occidente.”
3.9. Lobo a punta
|
sb
- spike blade |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Fu Warren (1939)
a
descrivere una mutazione della cresta semplice caratterizzata dal fatto di
terminare posteriormente in una punta singola invece dell’abituale lamina
larga e allungata. I soggetti non possono essere catalogati in questa
categoria fin quando non hanno raggiunto la maturitŕ sessuale. Pur essendo
una mutazione recessiva e non legata al sesso, le irregolaritŕ di
segregazione pare siano indicative del fatto che una piccola percentuale di
eterozigoti possa mostrare questo fenotipo, che talora viene soppresso in una
piccola percentuale di omozigoti. Le femmine esprimono questo tratto meglio
dei maschi.
3.10. Escrescenze laterali
|
Escrescenze
laterali |
Poligenico
Gruppo di associazione sconosciuto
Le
escrescenze laterali rappresentano una frequente anomalia della cresta
semplice ![]() . Considerate un difetto, molti allevatori risolvono il
problema tagliando il sovrappiů, ma in tal modo viene nascosto il fenotipo e
verranno sempre alla luce soggetti difettosi. Le escrescenze laterali
consistono in punte o dentelli extra, in numero da uno a tre, che nascono e si
sviluppano lateralmente nella parte posteriore della cresta semplice.
Abitualmente sono riconoscibili nei maschi all’etŕ di 3-4 settimane, mentre
le femmine debbono possedere parecchi mesi prima di essere accuratamente
classificate.
. Considerate un difetto, molti allevatori risolvono il
problema tagliando il sovrappiů, ma in tal modo viene nascosto il fenotipo e
verranno sempre alla luce soggetti difettosi. Le escrescenze laterali
consistono in punte o dentelli extra, in numero da uno a tre, che nascono e si
sviluppano lateralmente nella parte posteriore della cresta semplice.
Abitualmente sono riconoscibili nei maschi all’etŕ di 3-4 settimane, mentre
le femmine debbono possedere parecchi mesi prima di essere accuratamente
classificate.
Gli studi finora eseguiti hanno
dimostrato che non ci troviamo di fronte a un gene singolo. L’ipotesi
migliore č che si tratti di 2 geni autosomici dominanti complementari.
Punnett e Jull hanno ipotizzato una relazione tra questa anomalia e il gene del
ciuffo Cr.
3.11. Creste multiple
|
Creste
multiple |
Poligenico
Gruppo di associazione sconosciuto
I
tipi riscontrati da Taylor nel 1946 comprendono creste triple, quadruple e
quintuple, sempre a carico della cresta semplice. Dagli incroci effettuati si
č dedotto che sarebbe possibile creare una razza pura per questa
caratteristica, anche se il ceppo di Taylor non aveva ancora raggiunto uno
stato omozigote. Il 75% dei soggetti presenta una cresta quadrupla.
Dal momento che il ceppo di
partenza era dotato di escrescenze laterali, presenti anche in F1
e nei backcross, Taylor č dell’avviso
che sono coinvolti fattori multipli, alcuni dei quali con azione
complementare, alcuni dei quali senz’altro identici a quelli descritti da
Assmundson a proposito delle escrescenze laterali.
4. UROPIGIO
Uropigio
di gallo Livorno - Dopo averne asportato la cute che lo ricopre
si mettono in evidenza i due lobi che lo costituiscono.
Avian Anatomy Integument (1972) di Alfred Lucas & Peter Stettenheim
L’uropigio, o ghiandola dell’uropigio,
o ghiandola uropigetica, prende nome dal greco ourá,
coda, e pygaîon, deretano. Č una voluminosa ghiandola tubulare composta a
secrezione esterna costituita da due lobi, accolta nello spessore del derma
e posta dorsalmente alle ultime vertebre nella regione del codrione. Essa
elabora un secreto sebaceo che, dopo la confluenza del condotto ghiandolare
di destra e di sinistra, sfocia all’esterno attraverso un unico dotto
escretore terminale, il quale si apre in un rilievo della cute che prende il
nome di papilla.
Sezione
frontale di uropigio di gallo Livorno
Avian Anatomy Integument (1972) di Alfred Lucas & Peter Stettenheim
Il secreto dell’uropigio viene distribuito col becco a
tutte le penne, proteggendole cosě dall’essiccamento e soprattutto dall’umiditŕ.
Le molecole che lo costituiscono sono strutturate in modo tale da
incastrarsi l’una nell’altra mediante le loro catene laterali, per cui
le penne vengono ricoperte da una pellicola continua. L’uropigio manca in
uccelli che vivono in zone molto aride, č particolarmente sviluppato negli
uccelli acquatici, mentre in altri volatili ha dimensioni molto ridotte e in
tal caso la funzione protettiva viene assolta dal pulviscolo che si forma
quando le parti terminali delle penne si scompongono in finissime particelle
di cheratina.
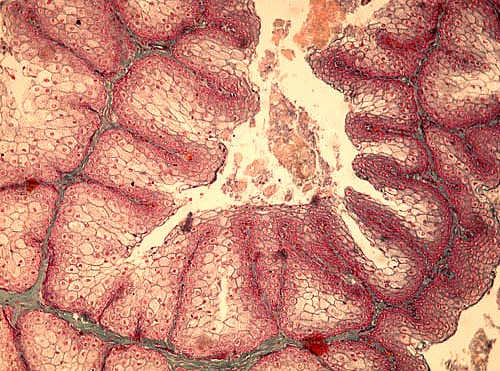
L'uropigio al microscopio ottico
A causa dei grassi che secerne, dal punto di vista
gastronomico l’uropigio rappresenta un boccone molto saporito anche nel
pollo arrosto. Vista la considerazione particolare in cui erano tenuti i
Ministri di Dio, gli venne attribuito il nome di Boccone del Prete. A
mio avviso suona invece un po’ irriverente il termine inglese riservato
all’uropigio: Parson’s nose, cioč naso del Parroco.
Quali sono le sostanze capaci di rendere cosě prelibato
questo boccone, detto anche cicerone?
Il termine cicerone non avrebbe nulla a che fare con l’oratore piů
famoso di tutti i tempi, mentre potrebbe derivare dall’accrescitivo del
latino cicer, che significa cece,
in quanto l’uropigio ha una certa somiglianza con un grosso cece. Ma anche
Cicerone![]() aveva a che fare col cece: infatti in origine molti nomi gentilizi
romani erano dei soprannomi presi dalle piante, tant’č che Fabio forse
deriva da faba, la fava, Lentulo
da lens, la lenticchia, e Cicerone
da cicer (Giuseppe Pittŕno, 1990).
aveva a che fare col cece: infatti in origine molti nomi gentilizi
romani erano dei soprannomi presi dalle piante, tant’č che Fabio forse
deriva da faba, la fava, Lentulo
da lens, la lenticchia, e Cicerone
da cicer (Giuseppe Pittŕno, 1990).
Ma torniamo ai componenti del secreto dell’uropigio.
Considerando i vari strati di cellule epiteliali dalle quali questa
ghiandola č tappezzata, essa č istologicamente simile all’epidermide. Le
gocce di lipidi cominciano a formarsi nelle cellule dello strato basale e
vanno progressivamente raggruppandosi tra loro, formando cosě delle
strutture globulari di dimensioni sempre maggiori. Tuttavia, siccome queste
cellule non vanno incontro a cheratinizzazione, esse vengono completamente
dissolte da parte dei lisosomi, verificandosi cosě una secrezione di tipo
olocrino. I principali costituenti idrofobici sono rappresentati da sostanze
ceruminose costituite da lunghe catene alifatiche e da omologhi complessi di
acidi grassi ramificati nonché da alcoli. Vengono prodotti lipidi sia
saponificabili che non saponificabili. Le specie acquatiche hanno ovviamente
bisogno di una quantitŕ maggiore di sebo, e la loro secrezione puň
contenere maggiori quantitŕ di triacilgliceroli.
I componenti chimici, oltre a proteggere la cheratina dai
fenomeni di sfaldamento, sono anche dotati di azione antimicrobica e
antifungina. Come accade per le ghiandole sebacee dei mammiferi, č
possibile che anche l’uropigio sia sotto controllo ormonale.
Le alterazioni gravi a carico di
questa ghiandola sono di scarsa importanza nei soggetti allevati in
batteria, mentre comportano l’eliminazione del ceppo se l’allevamento si
svolge al suolo e soprattutto all’aperto.
4.1. Sdoppiamento della papilla dell’uropigio
|
U
- uropygial |
Autosomico
incompletamente dominante
Gruppo di associazione I - cromosoma 2
Questa mutazione causa uno
sdoppiamento della papilla che č la
porzione anatomica dell’uropigio in cui confluiscono il condotto di destra e
di sinistra. Č stato Hutt (1949) a
fornire la descrizione degli effetti determinati dal genotipo. Gli eterozigoti mostrano un fenotipo che puň
andare da una
biforcazione completa della papilla sino a una perfetta normalitŕ dello sbocco ghiandolare. Parecchi omozigoti, alla nascita, presentano due abbozzi della
papilla, per poi andare incontro alla completa perdita della ghiandola, mentre
pochi possiedono una ghiandola sdoppiata con una papilla anch'essa doppia come
puň accadere agli eterozigoti. Č ovvio che l’espressione di questo gene č
condizionata da geni modificatori. Il locus U
č strettamente legato a quello della cresta a rosa.
4.2. Fissurazione e sdoppiamento della papilla dell'uropigio
|
U-2
- cleft and double uropygial papillae |
Autosomico
incompletamente dominante
Gruppo di associazione I - cromosoma 2
Questo allele del locus U
č stato studiato da Kessel (1945)
che
mise in evidenza come tutti i portatori di questa mutazione posseggano una
ghiandola funzionante, alcuni con una papilla sdoppiata, altri con una
fissurazione per tutta la lunghezza dello sbocco ghiandolare. Essendo spiccata
la differenza rispetto ai genotipi U,
Kessel ha proposto questa mutazione, ma potrebbe anche trattarsi del fatto che
la popolazione da lui studiata non segregasse per i geni modificatori come
accadeva per Hutt. Potrebbe pertanto trattarsi dello stesso gene.
4.3. Sdoppiamento della papilla dell’uropigio
|
dgp
- double oil gland papillae |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Mutazione studiata da Somes nel
1971, che puň causare da uno sdoppiamento completo della papilla a un solco
appena accennato alla sua sommitŕ. I test di linkage per il gene R hanno dimostrato una segregazione indipendente (51,5% di
crossover) per cui questa mutazione non risiede nel locus U.
5. TARSI E SPERONI
Lo
sviluppo di speroni č una caratteristica del maschio. Tuttavia, dopo un
giorno dalla schiusa, in ambedue i sessi se ne possono scorgere gli abbozzi,
formati da cheratina consistente. A circa 6 mesi d’etŕ lo sperone comincia
a ossificarsi, intorno ai 10 mesi d'etŕ si fonde
con l’osso del tarsometatarso attaccandosi cosě definitivamente allo
scheletro, e continua a crescere per tutta la vita.
5.1. Assenza di speroni
|
sl
- spurlessness |
Autosomico
recessivo, parzialmente limitato al sesso
Gruppo di associazione sconosciuto
La
mancanza dell’abbozzo epidermico dello sperone č giŕ documentabile al
primo giorno di vita, quando al suo posto si nota una larga squama cutanea.
Quest’anomalia genetica si accompagna spesso all’assenza di squame
epidermiche lungo la superficie interna dei tarsi. Abitualmente, raggiunta la
maturitŕ, si puň notare un dimorfismo sessuale, in quanto i maschi
manifestano lievi protuberanze nella sede degli speroni le quali successivamente si trasformano in escrescenze ossificate simili a speroni deformati,
mentre nelle femmine non si verifica nulla di tutto ciň.
Essendo un gene recessivo e parzialmente limitato a un
sesso, puň essere un’impresa alquanto ardua allontanare questo gene da un
ceppo in cui sia riuscito a penetrare, specie se si tratta di Combattenti, e
coloro che allevano il Combattente Inglese Antico debbono stare all’erta,
per non commettere l’errore di usare un riproduttore senza speroni. Se
malauguratamente sono incappati in questo disguido, faranno bene a utilizzare
come riproduttori solo i maschi dotati di speroni, tralasciando le loro
sorelle, delle quali č difficile conoscere il genotipo.
Non č descritto a quale gruppo di linkage appartenga
questo gene. L’unica osservazione personale consiste nel fatto che il mio Barbuto di Grubbe,
quindi senza coda, presenta anche un’assenza di speroni. Proviene dall’allevamento
del signor Rijs, in Olanda.
Chiesi a Mr Rijs se tutti i sui
maschi di Grubbe fossero privi di speroni. Se cosě fosse stato, si sarebbe
potuto pensare a un linkage tra l’assenza di posteriore e l’assenza di
speroni. Pochi giorni prima di ricevere la risposta da Rijs ne conoscevo giŕ il contenuto, in quanto
nel frattempo mi ero accorto che anche i miei maschi di Watermaal e
d’Anversa erano senza speroni. La lettera di Rijs č molto interessante, in
quanto, oltre a quesiti di genetica, suscita problemi estetici e filosofici.
Ingen,
03.04.1996
|
Dear Mr Corti, Referring to your question about the possible linkage
of the gene for spurlessness and rumplessness, I can give you the
following information: |
Caro Signor Corti Circa la sua domanda sulla
possibile associazione del gene per l’assenza di speroni e la mancanza
di posteriore, posso fornirle le seguenti informazioni: |
|
Spurlessness: A hot item these days in breeding Belgium bearded
bantam’s. Most of the cockerels and cock’s of the Antwerp, Watermaal,
Grubbe, Everberg & Uccle do not have spurs! According to an article
published in Germany in 1993, cocks without spurs are less or no
fertile. This based on research with commercial poultry. |
Assenza di speroni: Argomento scottante di questi
giorni tra gli allevatori delle Barbute Belghe Nane. La maggior parte
dei galletti e dei galli di Anversa, Watermaal, Grubbe, Everberg e Uccle non ha speroni! Secondo un articolo pubblicato in Germania nel
1993, i galli privi di speroni sono meno fertili o non lo sono affatto.
Questo dato č basato su ricerche fatte in linee commerciali. |
|
If this was the truth, we should not have such good
results in breeding Belgium bearded bantams because lack of spurs is
very common in these breeds. Anyway spurlessness is certainly not
reserved for the Grubbe Bantams. |
Se questo fosse vero, non
dovremmo avere cosě buoni risultati nell’allevamento delle Barbute
Belghe Nane, perché la mancanza di speroni č molto comune in queste
razze. Ad ogni modo, l’assenza di speroni non č certamente riservata
alla sola Nana di Grubbe. |
|
The general opinion in our country is that the
spurlessness is caused by the fact that we select our Belgium beardeds
very strongly on female related qualities. What I mean by that is that
we want absolutely no wattles, a very small comb, wide feathers and no
too much male-feathering. By breeding towards this direction we simply
forgot about spurs. |
Qui da noi l'opinione generale
č che la mancanza di speroni sia causata dal fatto che selezioniamo le
nostre Barbute Belghe puntando molto su caratteristiche che hanno
relazione con la femmina. Quello che voglio esprimere č che non
vogliamo assolutamente alcun bargiglio, una cresta molto piccola, penne
ampie e un piumaggio non troppo maschile. Quando incrociamo puntando in
questa direzione semplicemente ci dimentichiamo degli speroni. |
|
Since 1995 cocks without spurs are not allowed on
Dutch show’s anymore (cockerels are!). This rule was announced to the
breeders in 1990. Since that time breeders selected their breeding stock
on the gene for spurs. Result is on this moment that some of the male
birds do have their spurs back. |
In Olanda, dal 1995, i galli
senza speroni non sono piů ammessi alle mostre (sono galletti!). Gli
allevatori sono stati preavvisati di questa regola sin dal 1990. Da
allora gli allevatori hanno selezionato i loro ceppi in base al gene per
gli speroni. Il risultato č che in questo momento alcuni maschi hanno
di nuovo i loro speroni. |
|
In my case my black Grubbe do not have spurs but my
quail Grubbe do have spurs. We are aiming for spurs in our Watermaal and
Grubbe now. I noticed by experience that the surest way for getting back
spurs is breeding with females who came form a father who had spurs.
Some of their sons do have spurs even when the used father did not have
spurs. Breeding from a cock with spurs and females who came from a line
without spurs produced only males without spurs. Some other breeders had
the same experience to. But does this mean that the gene for spurs is
sex linked? |
Nel mio caso i miei Grubbe
neri non hanno speroni ma i Grubbe quaglia ne sono dotati. Attualmente
sto mirando a ottenere gli speroni nei miei Watermaal e Grubbe. Con l’esperienza
ho potuto notare che il modo piů sicuro per recuperare gli speroni sta
nell’incrociare con femmine che siano figlie di un padre con speroni.
Alcuni discendenti hanno speroni anche quando il padre impiegato in
riproduzione ne č privo. Allevando da un gallo con speroni e da femmine
provenienti da una linea che non li possiede, si ottengono maschi tutti
quanti senza speroni. Alcuni altri allevatori hanno avuto la stessa
esperienza in merito. Forse che il gene per gli speroni sia legato al
sesso? |
|
I hope that these informations do help you with your
research. |
Spero che queste informazioni
le siano di aiuto per le sue ricerche. |
|
Furthermore I like to share an experience with you
with is absolutely not clear to me: Each year we breed from our strain of cuckoo
Watermaal some chickens which are absolutely white (not yellow) in there
fluff. The rest of their brothers and sisters have the expected black
fluff with a white spot on their head. When these white chicks feather
they show a very light cuckoo colour with a lot of brown (rust) in it.
What can be causing this very strange colour? |
Vorrei inoltre farla partecipe
di un'esperienza che non mi č assolutamente chiara: Tutti gli anni dal mio ceppo
di Watermaal cuců escono alcuni pulcini dal piumino completamente
bianco (non giallo). I rimanenti fratelli e sorelle hanno l’atteso
piumino nero con la macchia bianca occipitale. Quando questi pulcini
bianchi mettono le penne, mostrano una colorazione cuců molto lieve con
parecchio marrone (ruggine). Cos’č che puň causare questa
colorazione molto strana? |
Per abbozzare una risposta a
Rijs sui soggetti cuců inabituali mi limito a darvi un’indicazione:
rivedere il barrato legato al sesso. Non aggiungo altro, in quanto vi priverei
del piacere di arrivare da soli a risolvere il problema. Ovviamente mancano
elementi di giudizio, in quanto Rijs non specifica il sesso dei pulcini, e l’ho
invitato a raccogliere dati precisi in tal senso, in quanto non č escluso che
abbia tra le mani la soluzione al dilemma se trattasi di un allele o di un’altra
mutazione che ha strette relazioni di linkage con il barrato legato al sesso.
Torniamo al problema degli
speroni. Giustamente Rijs fa notare che la fertilitŕ dei Barbuti non ha dato
problemi. Anche Veronica Mayew, in Inghilterra, non ha problemi di fertilitŕ
coi suoi Barbuti di Uccle, e non si era neppure accorta dell’assenza
di speroni nei suoi galli, in quanto le piume dei tarsi provvedono a
nascondere questa vergogna. Lo
studio tedesco si riferisce a polli commerciali, e spero di avere maggiori
notizie sia da Rijs che da Hoffmann, al quale mi sono rivolto per sapere cosa
ne pensa. Rijs afferma che i Barbuti sono fertili, perlomeno, lo sarebbero
come le altre razze, salvo differenze che possono sfuggire in un allevamento
amatoriale, ma comunque non tali da indurre a troncare l’allevamento di una
razza. Un’eventuale ridotta fertilitŕ nelle Barbute Belghe dovrebbe essere
causata da un’omozigosi per la cresta a rosa, ma la Barbuta di Uccle č
dotata di cresta semplice, per cui Rijs dovrebbe valutare, insieme agli altri
Soci del Club che si dedicano a questa razza, se la Barbuta di Uccle ha una
fertilitŕ maggiore rispetto alle altre Barbute, tenendo conto dei problemi d’accoppiamento
tipici della Grubbe e della Everberg.
Passiamo al problema filosofico.
Quali sono le caratteristiche che ci permettono di stabilire che un pollo č
un gallo e non č invece un cappone o una gallina? Il gallo canta, č
aggressivo, corteggia la femmina, si accoppia, ha la cresta e i bargigli, ha
gli speroni, ha piumaggio rosso e nero e sessualmente dimorfico. Sono poche le
razze che posseggono tutte queste caratteristiche. Pertanto, adottando il
concetto che i Barbuti senza speroni non hanno diritto di esibirsi alle
mostre, queste sarebbero piů desolate di quanto giŕ lo sono.
Sul canto del gallo non ho nulla
da obiettare. Il Gallo Rosso della giungla ha solo una piccola défaillance
durante la fisiologica fase d’eclissi, pronto a ridiventare gallo quando i
suoi ormoni sessuali glielo permetteranno. Il piumaggio bianco č contro
natura, in quanto non assicura la sopravvivenza, e lo sanno molto bene quegli
allevatori che debbono lottare contro falchi e sparvieri per assicurare una
boccata d’aria e d’erba ai loro polli. Gli Asiatici sono galli da poco, in
quanto a tutti č nota la scarsa aggressivitŕ che li caratterizza. Il
combattente Tailandese č praticamente senza cresta, per non parlare del gallo
Breda. I bargigli sono praticamente assenti nei Malesioidi, e tra loro
troviamo i galli combattenti piů robusti. Gli speroni sono un attributo
fisiologico anche di certe femmine che assicurano una buona progenie di galli
d’arena, pur avendo tutti i loro ormoni in ordine. Ai Barbuti gli speroni
non servono, e, come abbiamo selezionato il piumaggio bianco, nulla vieta di
selezionare galli senza speroni, che potrebbero anche presentarsi
esteticamente migliori. Il mio Barbuto di Grubbe, quello senza speroni, ha un’aggressivitŕ
da fare invidia a molti combattenti. Cosa dobbiamo pensare dei nostri galli travestiti?
Lord Sebright se la prenderebbe a morte
se sapesse che i suoi gioielli non sono considerati maschi. I combattenti con
piumaggio femminile non avrebbero diritto non solo alle mostre, soprattutto
non potrebbero calcare l’arena.
E dove li mettiamo i galli con
speroni multipli? In base alla filosofia olandese essi sono dei supergalli,
per cui bisogna riservare loro un padiglione apposito. Insomma, potremmo
andare all’infinito, e possiamo concludere che, se la teoria dell’out of
Africa č giusta, l’Uomo Bianco non č Uomo, in quanto č Uomo solo il
Negro.
5.2. Speroni multipli
|
M
- multiple spurs |
Autosomico
incompletamente dominante
Gruppo di associazione IV - cromosoma 2
Il Sumatra č l’unico a
possedere questa caratteristica distintiva di razza. I maschi maturi possono
portare da 3 a 5 speroni per zampa. Il secondo sperone, contato a partire dall’alto,
č il piů lungo; al di sopra si trova uno sperone piů corto, e, nel caso che
gli speroni siano 5, al di sotto del secondo si trovano altri 3 speroni brevi.
Ogni sperone ha un suo nucleo ossificato, ma solo i due superiori si attaccano
al tarsometatarso. Nei maschietti appena nati, come pure nelle femmine adulte,
il normale abbozzo unico č sostituito da 3-5 scaglie epidermiche allargate e
appiattite. Alla schiusa la diagnosi predittiva del numero di speroni presenti
nell’adulto puň essere fatta con un’accuratezza pari al 98%. A causa
della dominanza incompleta, gli eterozigoti presentano 2 speroni e gli
omozigoti 3-5. Alcuni soggetti, a causa di geni modificatori, qualora siano
eterozigoti, non presentano speroni multipli.
5.3. Sperone supplementare
|
As
- auxiliary spur |
Autosomico
dominante
Gruppo di associazione sconosciuto
Mutazione a penetranza completa,
caratterizzata dalla presenza bilaterale di sperone supplementare appena sopra
a quello normale. Questi speroni sono piccoli, a 18 mesi d’etŕ non superano
i 2 cm di lunghezza, si attaccano come di norma al tarsometatarso, e si
allungano col passare del tempo. Solo occasionalmente si riscontra un secondo
sperone soprannumerario appena al di sotto di quello normale. Il momento
migliore per diagnosticare quest’anomalia č a 20-40 settimane, non alla
nascita. Nel caso di un terzo sperone, č stata suggerita la possibilitŕ di
un parziale effetto dose del gene As
presente allo stato omozigote.
5.4. Sperone doppio
|
ds
- double spur |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Domm (1931)
ha
riferito della presenza di un doppio sperone in 3 femmine Livorno dopo essere
state castrate, e volle attribuirla a una mutazione genetica. Piů tardi
Warren ha studiato dei pulcini con sperone doppio, dimostrando che si trattava
di uno sdoppiamento giŕ a carico dell’abbozzo, presente abitualmente in
ambedue i tarsi e talora in uno solo. Raggiunta la maturitŕ, gli speroni si
svilupparono distinti l’uno dall’altro, in quanto la fissurazione giungeva
a interessare il nucleo basale. Talora le dimensioni degli speroni erano
diverse da una zampa all’altra. Warren fu pure in grado di stabilire che l’ereditarietŕ
di questo tratto č influenzata da fattori sconosciuti. Anche se autosomico e
recessivo, ds manifesta la sua
azione piů spesso nelle femmine, con scarsa penetranza negli omozigoti.
5.5. Muta degli speroni
|
Muta
degli speroni |
Ereditarietŕ
sconosciuta
Nel 1983 Kimball ebbe la fortuna
di descrivere un gallo che annualmente perdeva gli speroni in occasione dell’abituale
muta. Un esame ai raggi X dimostrň che essi non si saldavano mai all’osso
della zampa. Non č stato possibile proseguire in studi genetici, ma
probabilmente il comportamento di questa caratteristica era di tipo recessivo,
visto che nessun maschio di F1 e F2
mostrň quest’anomalia.
5.6. Galline con speroni
|
Galline
con speroni |
Ereditarietŕ
sconosciuta
Č piuttosto raro trovare
speroni in femmine di razze pesanti, mentre č frequente il loro riscontro
nelle razze mediterranee e nell’Amburgo. Per l’analisi di questo argomento
si rimanda al capitolo dedicato ai caratteri sessuali secondari. Si puň
aggiungere che, secondo l’esperienza di Somes, e secondo quanto riferito da
un allevatore statunitense di Galli d’arena le cui femmine erano dotate di
speroni, si puň concludere che, quando galline con speroni vengono incrociate
con razze leggere (Livorno, Polish, Sebright), parecchie figlie sviluppano
speroni. Č necessario procedere a uno studio genetico approfondito per dare
una risposta esauriente.
Dal punto di vista storico rammento che gli speroni nella gallina sono stati citati e disapprovati sia da Plinio (X,156) che da Columella (VIII,2,8) e che ne abbiamo parlato in capitoli precedenti.
5.7. Sindattilia
|
Sindattilia |
Effetto
pleiotropico di psp?
Implicati i geni dell’impiumamento dei tarsi?
In
questa condizione anatomica si ha la fusione fra 3° e 4° dito, per lo piů
attraverso una membrana interdigitale ininterrotta la cui estensione varia
da caso a caso e che nei gradi estremi comporta una stretta unione delle
dita tra loro, con l’unghia del 4° dito deviata rispetto alla sua normale
posizione, rendendo la deambulazione difficoltosa.
Questo tratto si esprime
abitualmente in ambedue le zampe. Circa le modalitŕ di trasmissione,
esistono numerose ipotesi, tra cui troviamo quella secondo cui l’associazione
con brachidattilia (By) e ptilopodia (Pti-1
- Pti-2 - pti-3) sarebbe dovuta a uno stesso gene, la cui azione viene
modificata dai soliti ignoti,
cioč da geni modificatori. C’č chi ha espresso per questo tratto un
comportamento dominante, esiste anche l’ipotesi che la responsabilitŕ
ricada su due geni dell’impiumamento dei tarsi. Da ricordare infine che č
stata proposta l’azione di un singolo gene pleiotropico psp,
descritto nella parte generale a proposito della pleiotropia (vol.II
- XIV.6). Da quanto detto, risulta lampante che la scienza ne ha
fatta di strada, ma che la soluzione di certi interrogativi č rimandata
di alcuni decenni. Molto verosimilmente esiste piů di un tipo di
sindattilia, e quello che pare piů certo č che essa non č legata al
sesso.
5.8. Dattilolisi
|
dac
- dactylolysis |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Č
una sclerodermia - dal greco sklërós+dérma = dura pelle - che
colpisce i piedi dei giovani pulcini, i quali sono normali all’atto della
nascita, ma dopo una settimana cominciano a presentare in tale sede un
irrigidimento della cute associato ad aspetto vitreo per un lieve edema. In
pochi giorni si instaurano delle fissurazioni alla pianta dei piedi che
possono estendersi anche dorsalmente. L’apice delle dita č atrofico e
assottigliato. In alcuni soggetti questa situazione non č grave e si
risolve nel giro di alcune settimane, mentre nei casi piů severi furono
necessarie anche 12 settimane per una restitutio
ad integrum.
Ricorrendo agli incroci si č potuto dimostrare che quest’affezione
č ereditabile e non č da attribuire a disordini alimentari. Puň essere
classificata tra le condizioni semiletali per le difficoltŕ che causa nell’affrontare
i primi atti della vita. Sotto certi aspetti č simile alle lesioni
provocate da deficienza di biotina oppure di acido pantotenico, talora
assomiglia alle vesciche ai piedi
che tra poco descriveremo.
La
biotina, detta anche vitamina H o vitamina antiseborroica, viene
sintetizzata dalla flora batterica intestinale,
per cui č difficile si instauri una sua carenza da scarso apporto, mentre
nell’uomo puň determinarsi una sua mancanza per un eccessivo consumo di
albume crudo. Infatti il bianco dell’uovo contiene l’avidina, che si
combina con la biotina a formare un composto non assimilabile. I sintomi
principali dell’avitaminosi H consistono in cute secca, anemia e
ipercolesterolemia.
L’acido pantotenico č anche detto fattore
antipellagra del pulcino e la sua carenza non dŕ segni nell’uomo. Č
contenuto nel fegato, nel rene, nel lievito e nella verdura fresca. Se i
pulcini non dispongono di questa vitamina in dosi adeguate, essi presentano
una dermatite, o pelle agra, cioč
una pelle ruvida.
5.9. Vesciche ai piedi letale
|
bf
- blistered foot lethal |
Autosomico
recessivo
Gruppo di associazione sconosciuto
Somes
(1970) ha riportato un’anomalia del piumino che si presenta accorciato, cui
si associano lesioni della faccia e dei piedi. Queste alterazioni sembrano
tutte dovute all’azione di un solo gene, spesso letale. Su 58 pulcini
vivaci e vitali all’atto della schiusa, il 25% morě al primo giorno, e di
quelli rimasti il 96% morě in capo a 18 giorni. Solo due sopravvissero fino
all’etŕ adulta, senza perň raggiungere la maturitŕ sessuale. Le
vesciche alla pianta dei piedi erano presenti all’atto della schiusa, e in
coloro che sopravvissero piů di 8 giorni si sviluppň una dermatite
ingravescente cui fece seguito una necrosi cutanea con perdita delle dita e
successive lesioni agli angoli della bocca e agli occhi. Nei due
sopravvissuti le lesioni si estesero anche alle orecchie. Coda e ali erano
iposviluppate.